博文
氢气生物活性的理论评估(博士之4)
||
读本文先阅读:
第四章:氢气生物活性的理论评估
4.1 引言
氢气是由两个氢(H)原子结合而成的,具有107 kcal/mol(4.64 eV)的H-H键能(Kim等人,2022)和相对于标准氢电极的还原电位(2H + 2e- → H2)为-0.421 eV(pH 7)(Karp, 2008)。尽管在生物系统中的主要作用方式尚未阐明,但一种受青睐的假设是氢气能够通过与自由基和离子物种(例如•OH、ONOO-)直接相互作用来减少过量的ROS/RNS。然而,由于氢气的空间和时间可用性,反应动力学在体内是否有利仍然是个争议点(Penders等人,2014;Hancock, LeBaron和Russell, 2021;Li等人,2021)。此外,虽然氢气治疗的临床优势现已得到充分记录(Korovljev等人,2017;Guan等人,2020;Botek等人,2022;Kura等人,2022;Tao等人,2022;Deryugina等人,2023;Wang等人,2023a),但关于氢气的分布及其与生物活动相关的精确分子机制仍有许多问题。这些问题包括:氢气如何到达目标组织?氢气相互作用的主要生理目标是什么?以及,氢气如何随时间保持其影响?
随着越来越多的动力,国际研究机构正在投入时间和资源来了解氢气如何影响细胞和更广泛的体细胞生理学。本报告评估了围绕氢气分布和分子活动的初步研究,并就氢气在生物系统中的主要作用模式进行了理论化。
4.2 还原活性气体
4.2.1 羟基自由基
•OH被视为所有ROS中对生理最有害的,部分原因是因为氧化细胞分子可以迅速发生。在细胞应激事件中,超氧阴离子的增加产生是通过NADP+/H和黄嘌呤氧化酶(XOs)等产生ROS/RNS的酶的还原反应(Halliwell, 1978),或通过呼吸电子传递链(ETC)中的功能障碍过程(Ratajczak等人,2019)发生的。后者是由于FMN和Q模块的复合物I(第3章,第3.6节)以及复合物III(辅酶Q-细胞色素c氧化还原酶)中的O2过早还原导致电子泄漏的结果。酶还可以产生超氧阴离子,无论是作为对抗病原体攻击的防御性氧化爆发,还是作为线粒体功能障碍的后果(Wang等人,2018b)。在这种情况下,超氧阴离子的产生利用Fe2+或Cu2+金属蛋白催化O2的单价还原,电子由NADH(2O2 + 2e-)或NADPH辅因子捐赠。超氧阴离子的歧化速率常数为8 x 10^4 M^-1 sec^-1(pH 7.8),尽管这是二阶速率反应(与反应物浓度的平方成正比)。一阶速率反应(反应速率直接与单一反应物的浓度成正比)被计算为快106倍。在没有清除剂的情况下,超氧阴离子在生物系统中的半衰期从0.05秒(超氧阴离子浓度:1 x 10^-4 M)到14小时(超氧阴离子浓度:1 x 10^-10 M)不等(Fridovich, 1983;Gao等人,2007),见表4.1。
表4.1. 细胞中ROS/RNS活动的物理限制。数据摘自并改编自Zhang等人(2019a)。寿命:自然对数2除以所有ROS反应的分子的速率常数和浓度乘积之和。扩散距离:使用公式x = (6Dt)^(1/2)计算(x、D、t分别代表扩散距离、扩散系数和寿命)。
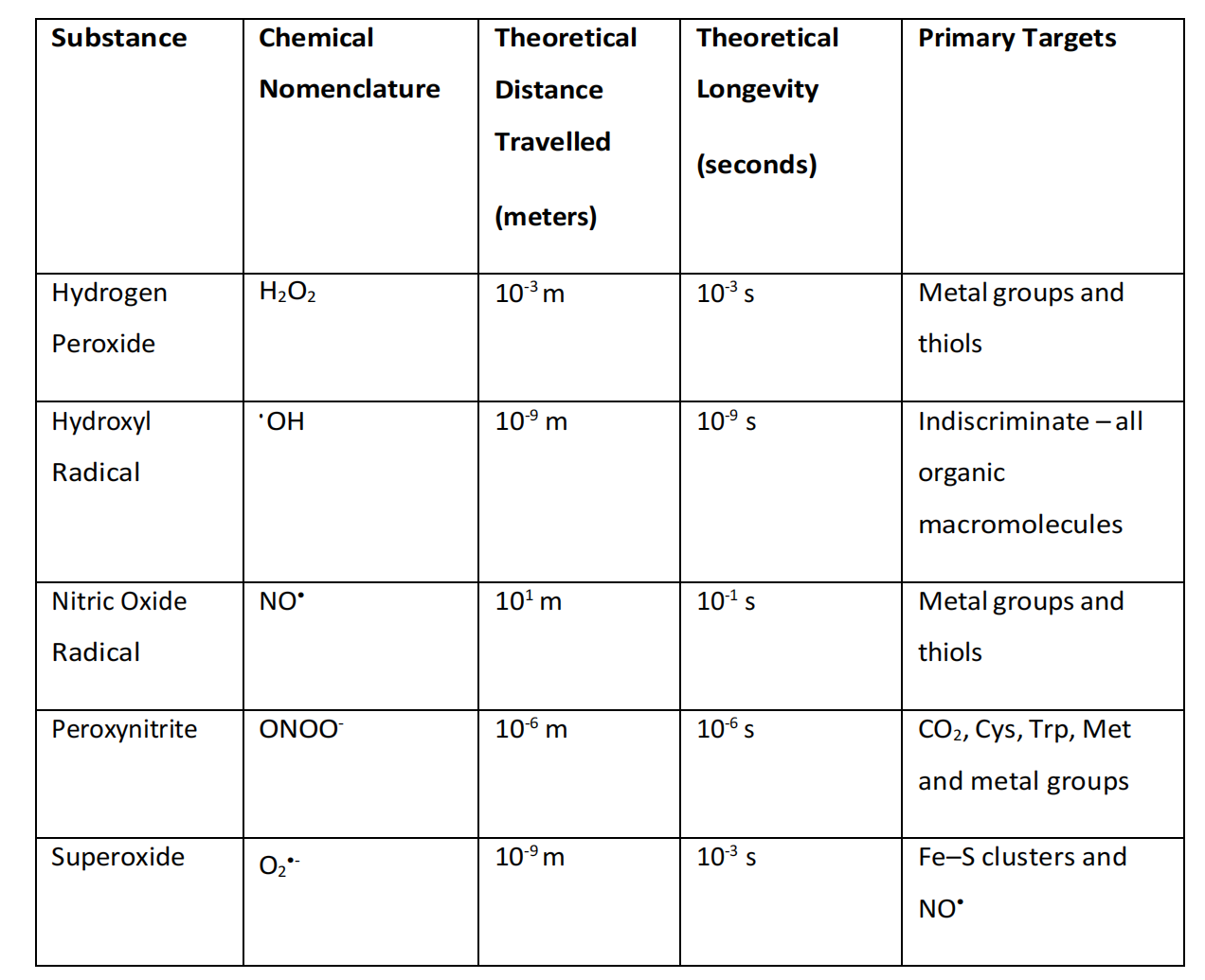
超氧阴离子被视为最基础的ROS,因为其形成已知会导致ROS和RNS的产生,包括•OH。在这里,通过SOD的歧化作用,超氧阴离子产生H2O2,后者可以通过下面提到的下游反应生成•OH。
•OH是通过分子氧的三电子还原形成的(Di Meo和Vindetti, 2020),已知生物体内羟基自由基的形成有多种方式。例如,Fenton反应和Haber-Weiss反应在H2O2存在下都会产生•OH。
Fenton反应:O2•- + H2O2 → O2 + •OH + OH-(在铁离子存在下)
Haber-Weiss反应:Fe2+ + H2O2 → Fe3+ + •OH + OH-
超氧阴离子的还原可能是由内源性SOD活性引起的,或者是通过Fenton/Haber-Weiss反应,尽管后者估计在细胞中产生的H2O2水平相对较少(Winterbourn, 2020)。已知超氧阴离子的还原是H2O2的主要来源,H2O2是一种能够穿越磷脂膜并影响细胞和细胞器活动的信号分子(Krohn, Maier和Paschke, 2007; Iglesias-Pedraz和Comai, 2020)。静电吸引将超氧阴离子吸引到包括各种铁氧还蛋白、还原酶和脱氢酶在内的酶的暴露铁硫簇上,可以通过释放Fe2+/H2O2阵列来生成H2O2(Demidchik, 2017)。释放Fe2+/H2O2阵列可以影响对酶硫醇的翻译后修饰,激活或失活蛋白质活性,这种效应具有许多但较少明确定义的下游信号效应。这种蛋白质修饰的反应速率计算为>106 M-1 s-1(Winterbourn, 2020)。
Kim等人(2022)对氢气在血红素蛋白中的预期效果进行了开创性计算,说明了通过氢气与原血红素之间的相互作用实现抗氧化活性的可行性。作者报告说,氢气分子可以通过二氢键不对称地结合到血红素中的铁(Fe2+)上,并通过称为Kubas键合的双侧电子转移对称结合。在生理条件下,由于激活能相对较低(分别为2.04 eV和2.14 eV),不对称结合被认为是更有利的。作为回报,Fe2+应该降低氢气二聚体中H-H键的解离能,形成Fe2+/H•复合物,其中H•能够还原高活性的离子和自由基。Fe2+/H•复合物的解离能相对较低,为2.78 eV,这将允许结合的H•自由基中和另一个共时的活性硝化或氧化物种(Kim等人,2022)。
因此,作者声明这种提议的活动可能解释了•OH的直接还原,然而这种提议的作用模式的动力学尚未阐明。•OH也可以通过ONOOH的均裂产生,其在25°C下的衰减速率常数为1.2−1.3 M-1 s-1,生成•OH和二氧化氮(NO2•)自由基(Merényi和Lind, 1998)。已知•OH以扩散控制速率与生物系统中的•OH反应(1 x 109 M-1 s-1)(Halliwell等人,2022; Lenzen, Lushchak和Scholz, 2022),表明这个分子在与生物化合物反应之前无法传播很远。Li等人(2019)建议•OH的作用将限于其生成地点的4.5 - 8.6nM范围内(Li等人,2019; Zhang等人,2019a)。由于•OH可以在细胞内由一系列酶(例如,脱氢酶、水合酶、还原酶、合成酶)全局产生,氢气需要在•OH产生的精确位置可用,并且数量显著大,以有助于直接清除•OH。当相对较小的氢气剂量被证明具有治疗效果时(Alharbi等人,2022),这是不太可能的。
支持氢气不太可能对•OH的直接还原有显著影响的假设,•OH与氢气的生物分子反应动力学计算为4.2 x107 M-1 s-1(Li等人,2020),与•OH和包括脂质和蛋白质在内的大分子的氧化反应相比,活性明显下降(> 1x 109 M-1 s-1),至少比氢气引发的任何潜在还原高出两个数量级。除了氧化大分子外,•OH还通过氧化活动影响氨基酸化学,其动力学也应该超过氢气的还原潜力。氨基酸如脯氨酸和谷氨酸与•OH的反应速率常数>2.3 x 108 M-1 s-1(Ishibashi, 2019),而与谷胱甘肽的反应计算高达>1x1010 M-1 s-1(Tao, Song和Qin, 2019)。这表明氢气直接有效地清除生物系统中的•OH可能是微不足道的。因此,尽管被提议为一种有效的抗氧化剂,具有潜在的作用模式,如Kim等人(2022)所报告,但氢气直接有效清除氨基酸、气体、脂质等生物环境中的•OH的能力令人质疑。
总结来说,•OH作为一种自由基物种,可以与大多数有机分子(包括脂质、核苷酸、肽和蛋白质)反应,因此更有可能与局部的生物成分相互作用,而不是与氢气。考虑到上述不利的动力学因素,以及高反应性、短半衰期和这类活性物种的最大扩散限制,氢气通过主要作为清除剂来清除生物系统中的•OH而发挥广泛的抗氧化作用是不太可能的。
4.2.2 过氧亚硝酸离子
现在已充分确定,NO•作为二级信令分子,在生物化学中具有多种作用。已知NO•既能激活抗氧化转录途径,在有氧条件下也能与含氧自由基物种反应,形成更稳定的硝酸钾和亚硝酸钾产物(Hancock和Veal, 2021)。气体传递物质如NO•的活动通常取决于它们的细胞浓度,浓度超过生物学阈值可导致细胞毒性过程的诱导,这些过程涉及自由基级联的启动,以及对蛋白质的氧化和亚硝化翻译后修饰(Del Castello等人,2019;Hancock和Veal, 2021)。NO•的毒性主要归因于下游氧化产物,包括顺磁性氮氧化物NO2•和ONOO-。
当NO遇到超氧阴离子时,它可以自发反应,形成ONOO-,其动力学大于内源性抗氧化剂SOD(分别为6.75 x 10^9 M^-1 s^-1 vs 6.4 x 10^9 M^-1 s^-1)(Speckmann等人,2016)。因此,ONOO-的自发产生可能比超氧阴离子的歧化成O2和H2O2更快。
ONOO-是一种高度反应性和可扩散的分子(表4.1)。这些特性以及在实验条件下避免氧气的需要,意味着H2/ONOO-相互作用的速率常数尚未得到确认。然而,在少数研究H2/ONOO-反应性的研究中,Penders等人(2014)指出过氧亚硝酸(ONOOH,ONOO-的质子化形式)的降解产物没有显著增加,并确定在没有ONOO-/大分子相互作用的情况下自发衰减速率为1.1 - 1.3 M^-1 s^-1,无论是否应用氢气(Ferrer-Sueta和Radi, 2009;Penders, Kissner和Koppenol, 2014)。ONOOH在这里很重要,因为估计其pKa为6.6,表示弱酸,生物系统中可能存在生物学相关的ONOOH量(Villamena, 2013)。当检查这些氧化还原伙伴的下游效应时,这也很可能是重要的。ONOOH均裂的产物是NO2•和•OH(2.5 M^-1 s^-1),而ONOO-离子已知会产生超氧阴离子和NO•(0.017 M^-1 s^-1)(Merényi和Lind, 1998)。在生理系统中,ONOO-还与二氧化碳(CO2)反应,速率常数为5.8 x 10^4 M^-1 s^-1,形成三碳酸(CO3•-)和NO2•作为分解产物(Squadrito和Pryor, 2002),这些都是已知会影响众多细胞反应的反应性物种。
在使用选定的离子流管/质谱法研究ONOO-和氢气之间的反应动力学时,没有显示出这些物种之间的反应(Viggiano, Midey和Ehlerding, 2006)。因此,可以推测氢气和ONOO-的反应动力学,在温度和pH等生理参数下尚未确定。尽管如此,目前的理解是氢气导致ONOO-水平选择性降低,和/或在生物系统中减少过氧化损伤。事实上,Kiyoi等人(2020)描述了在小鼠模型中通过氢气吸入治疗时•OH和ONOO-产生的减少。有趣的是,同一研究(Kiyoi等人,2020)指出,负责在细胞应激事件中产生超氧阴离子的NADPH氧化酶(NOX-1)酶在氢气组中显著下调。关键激活组件p40-phox和p47-phox的表达不受影响,表明氢气可能影响NOX-1复合物内其他蛋白质的表达或作用。NOX酶促进O2转化为超氧阴离子,利用Fe+2/3过渡金属作为电子转移的催化剂,因此可能为氢气相互作用提供一个可行的目标(Hancock, LeBaron和Russell, 2021),尽管需要实证数据来确认这一点。
4.3 氢气的分布和保留
4.3.1 药代动力学
对猪模型中氢气分布的药代动力学分析(Sano等人,2020)表明,由于氢气在静脉血中的半衰期增加,这种二原子气体可能通过血浆传播,并简单地扩散到组织结构中。氢气的物理和化学特性(第1章,第1.1节)应允许该分子通过生物液体、细胞外基质、细胞膜和胞质区室扩散。然而,这种分散机制是否能够解释在远端器官中看到的效果尚不确定。
例如,当通过腹膜内给药给予富氢盐水(HRS)时,记录到胰腺和脾脏等近端部位有大量氢气,而远端器官则显示逐步减少,呈放射状进展(Liu等人,2014)。有趣的是,当通过吸入给药氢气时,虽然主要受益器官通常靠近(例如,大脑、呼吸系统和心脏),但在给药后30分钟也看到脾脏和骨骼肌中氢气浓度显著增加。如果氢气暂时驻留在血红蛋白的微孔或口袋内,或者血清中的悬浮分子如碳水化合物或无机离子(例如,糖原和钙)暂时保留氢气,就可以解释这个因素;在第4章,第4.3.2 – 4.3.4节中讨论了这一点(Diamantis, Unke和Meuwly, 2017;Hancock等人,2022;Hancock, 2023);
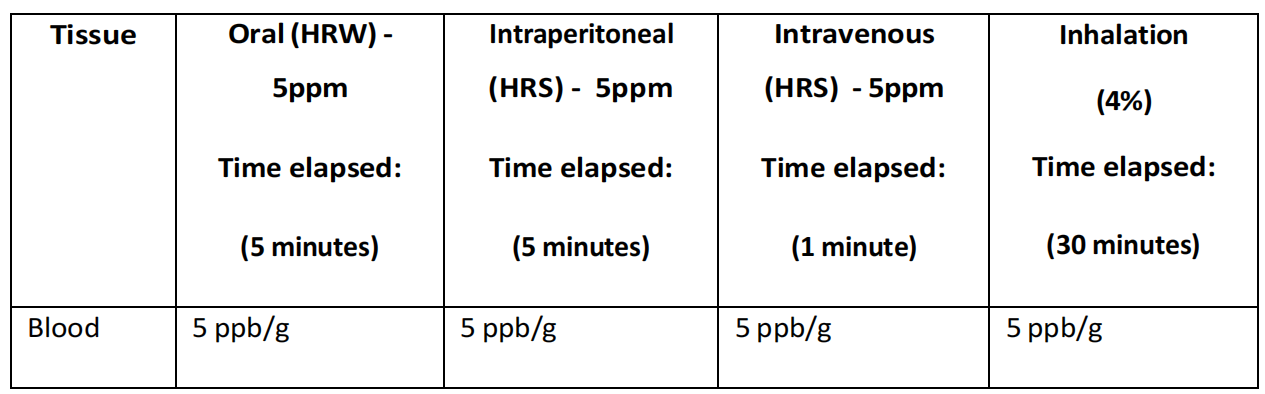
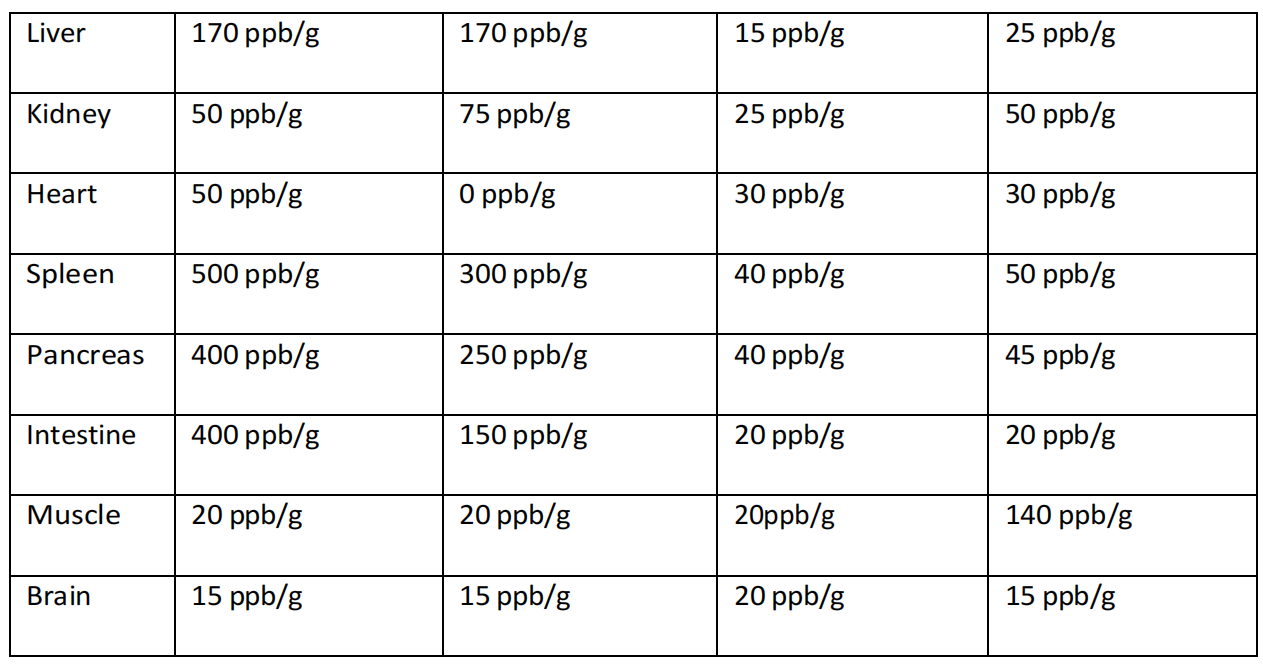
尽管这项研究(Liu等人,2014年)发现通过腹腔注射富氢盐水(HRS)可以在分析的大多数组织中提高氢气水平,但这种方法在将大量氢气输送到血液中或更广泛的心血管系统中并不有效,而在将氢气输送到近端器官,包括肠道、肝脏、胰腺和脾脏时却非常有效(表4.2)。这些结果表明,分布更倾向于简单的扩散方式,然而,这仍然无法解释在未检测到血液中氢气水平升高的情况下,远端器官如大脑和肾脏中氢气水平升高的现象(表4.3)。
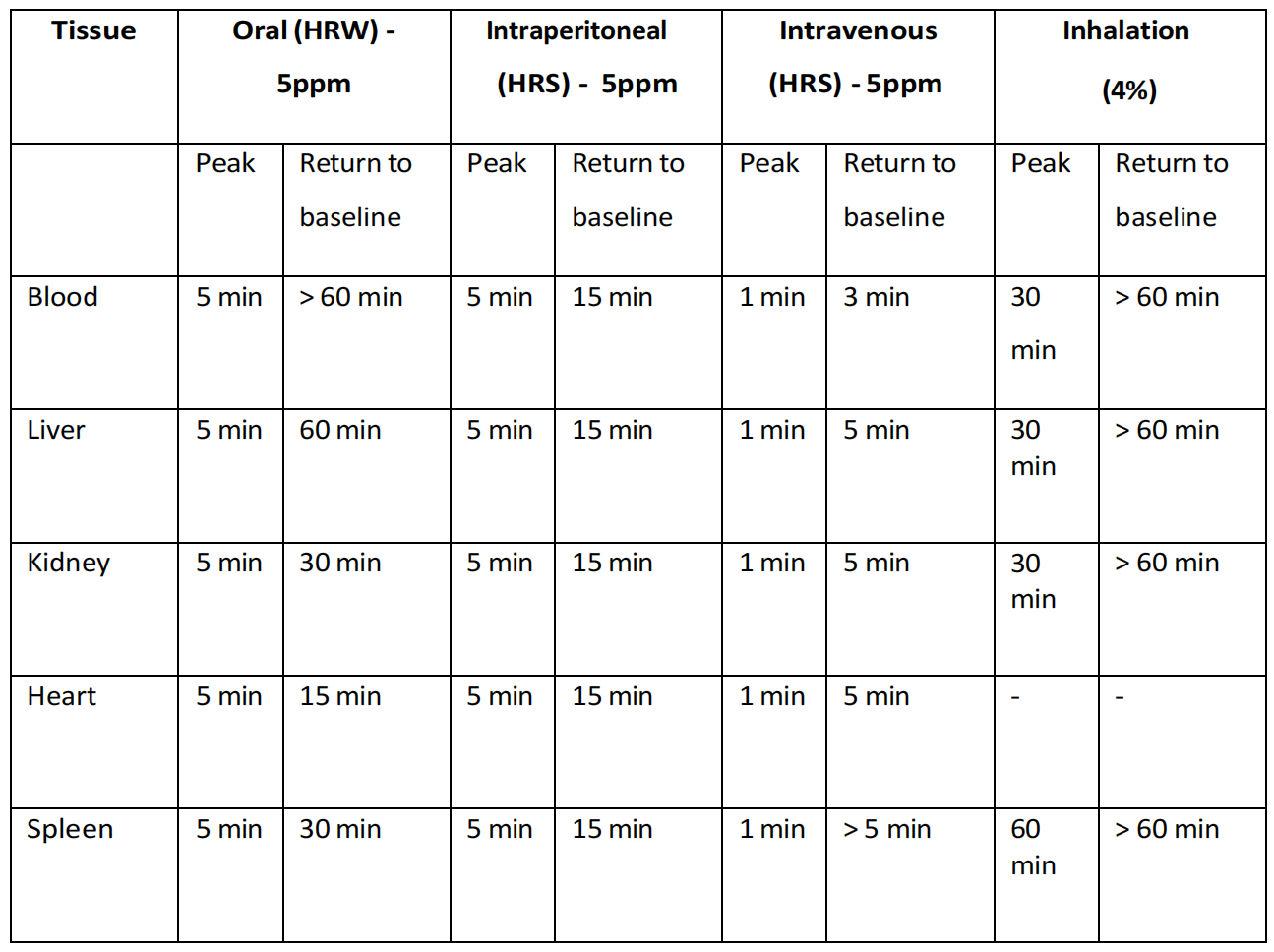
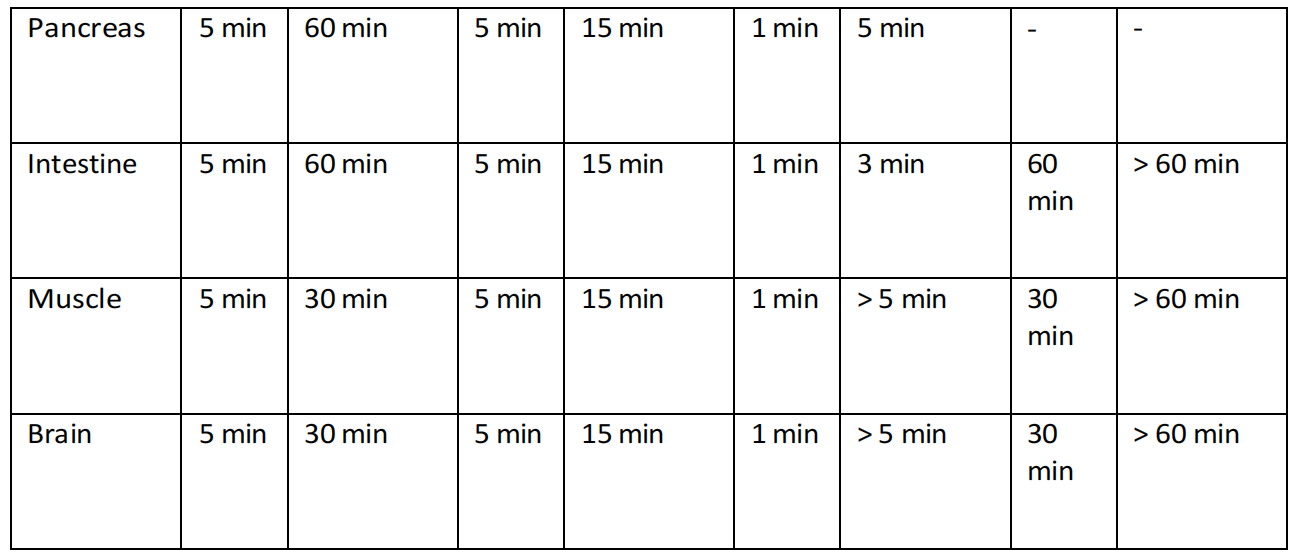
这种现象的一个原因可能是因为实验协议只记录了溶解在血浆中的氢气。血浆的饱和点在1.6mg/L/氢气附近,因此吸入可能会提供更多的氢气,超过血液能够溶解的量。那么问题来了,足够的氢气是如何通过血液传播的,以至于在治疗后长达一小时的时间里,静脉血液中和经常是远端器官中仍然可以检测到氢气的存在?
一项研究调查了100% 氢气吹入(将某物吹入体腔的行为)的效果,得出结论认为氢气很可能留在血浆中(Christ等人,1994年)。然而,要出现这种情况,饱和水平必须大大增加,才能看到如此广泛的影响和分布模式。由于生理压力仅比大气压(760 mm/Hg)高出约120 mm/Hg,因此由于压力增加而导致氢气的饱和水平升高是不可能的。此外,通过血浆的简单扩散无法解释骨骼肌中氢气逐渐增加和保留的情况,也无法解释为什么氢气不仅在输送部位附近的器官有效。然而,氢气与葡萄糖分子或金属蛋白(例如血红蛋白和肌红蛋白)的结合,可能为了解氢气如何在躯体系统中分布、保留和/或利用提供了一种见解。
4.3.2 血红素
血红素是一种含铁的辅基团,存在于所有脊椎动物物种的细胞色素和红细胞中,除了南极冰鱼(Channichthyidae)(Sidell和O’Brien, 2006年)。血红蛋白蛋白质在生物气体(例如,O2、NO、CO和CO2)的运输和交换中起着主要作用,通过直接与血红素辅基团的相互作用以及对硫醇和胺部分的共价修饰(Hancock等人,2022年),由4个单独的亚基组成,每个亚基都能结合一个O2分子。在血红蛋白(和肌红蛋白)中,Fe2+在N-卟啉环内五配位,第六位置结合氧气(图4.1)。
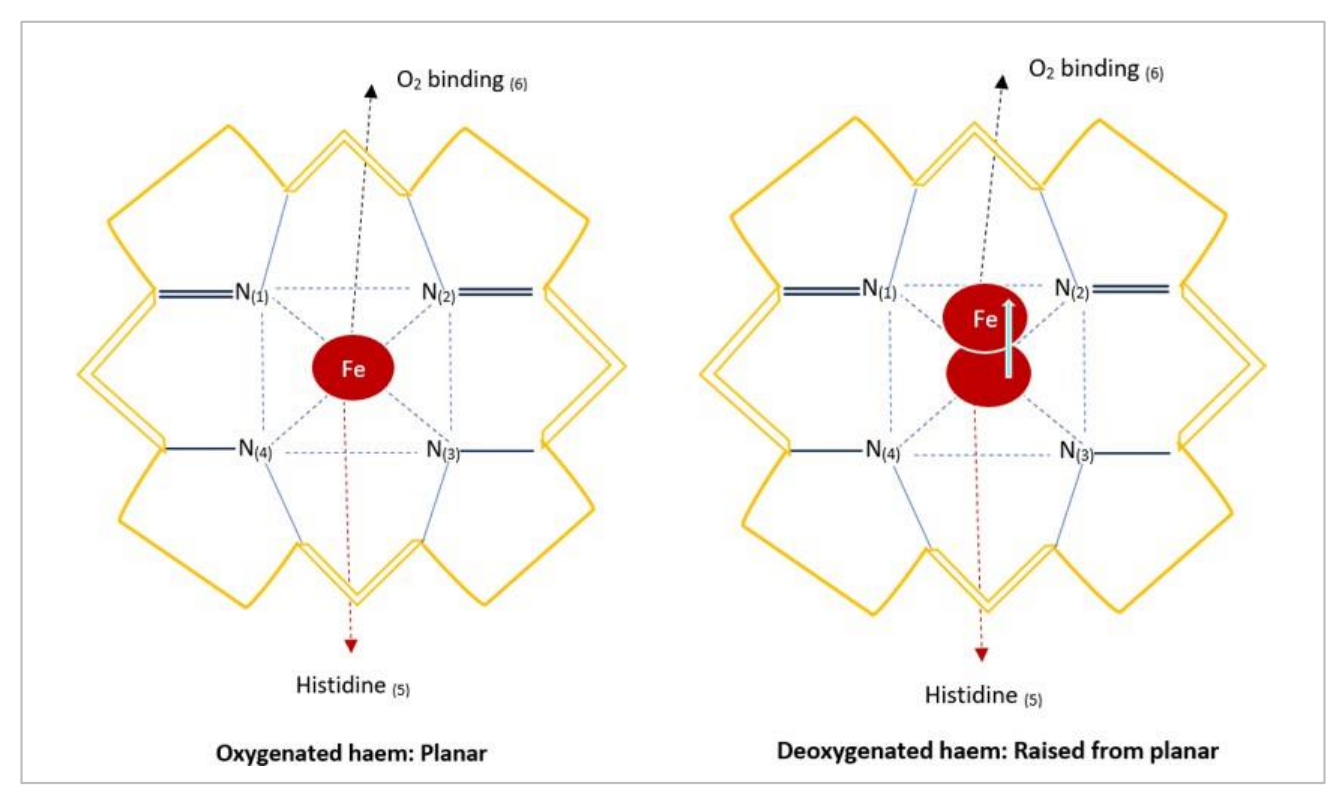
图4.1. Fe2+血红素辅基团的图形表示。左侧 - 氧合血红素(松弛状态)。右侧 - 脱氧血红素(紧张状态),Fe与卟啉环的距离增加了0.06nm。括号指示结合位置。His – 代表蛋白质的Fe+2/3结合组氨酸残基(例如,血红蛋白)。
血红素具有多功能性,可以以多种状态存在,每种形式的含量都会影响生理过程。例如,亚铁离子(Fe2+)存在于脱氧(脱氧-)血红素和氧合(氧合-)血红素中,而铁离子(Fe3+)存在于(甲-)血红素中,这是一种非氧结合构型。此外,与Fe的离子状态相关的血红蛋白的构象有显著差异。例如,脱氧血红蛋白被称为“紧张”状态,因为球蛋白从卟啉环向远端移动了0.06nm(Heremans, 1987)。另一方面,氧合血红蛋白被定义为处于放松状态,球蛋白分子更靠近卟啉环(图4.1)。氧化到Fe3+(甲-血红素)的变化导致血红素口袋的几何形状发生变化,有利于放松位置,使得氧气不再能够结合。
血红素的铁磁性也可能影响气体的结合特性(Hancock和Hancock, 2018)。例如,通过硅片分析亚铁离子的自旋态发现,Fe-N键距离从低自旋态的2.01Å增加到高自旋态的2.1Å(Ugalde等人,2004年)。因此,由于高自旋态的Fe2+可能从卟啉环延伸得更远,高自旋态的Fe2+在吸引O2(和氢气)到血红素辅基上可能比低自旋态的Fe2+更有效。
当研究氢气对血红素构型和效用的影响时,注意到每日给予HRW(8mM/L/天)可以提高中年女性的O2饱和度(Ostojic等人,2018年)和COVID-19康复患者的O2饱和度(Singh等人,2021年)。为了提出这种效应可能发生的原因,并考虑到血红素辅基团的松弛和紧张状态,氢气可以通过两种方式影响血红素的氧化。首先,氢气可以通过双边Kubas键合(涉及两个氢原子同时向铁中心捐赠电子)保护Fe2+不被氧化,其中Fe2+/H2排列可以直接减少•OH自由基(例如,H2+2 •OH → 2 H2O)和/或其他活性物种(Kim等人,2022年)(图4.2)。其次,如果高自旋态的Fe2+可以增加Fe2+ -N键长,那么氢气有可能通过延长2.0Å的Fe2+ -N键改善氧附着到低自旋亚铁的血红素辅基上(图4.3)。
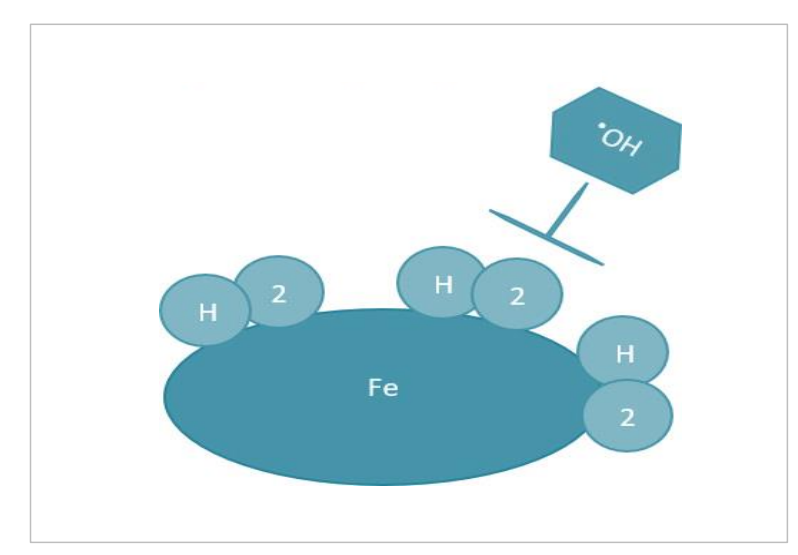
图4.2. 防止Fe氧化的氢气可能机制。示意图展示了氢气通过Kubas键合到Fe上可能防止氧化的过程。
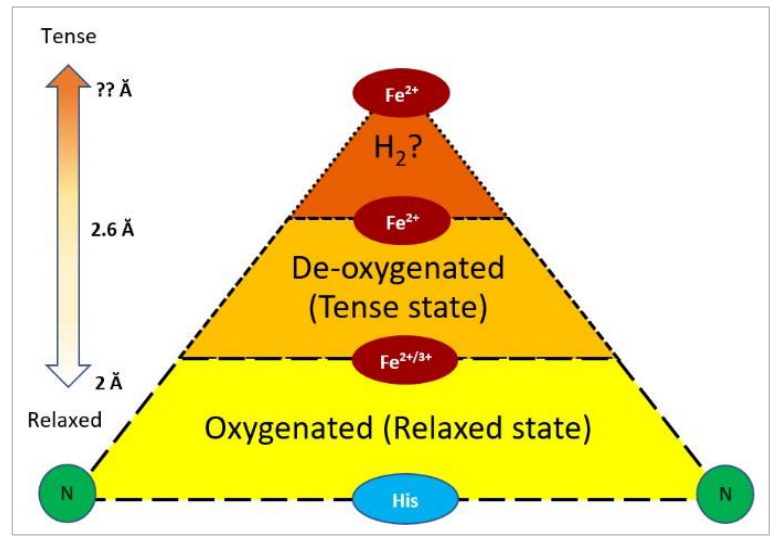
图4.3. 血红素跳板。单一方面的示意图表示在N-卟啉环和铁辅基团之间的不同键长。底部(黄色):代表氧合血红素的较短键。中间(橙色):代表脱氧血红素的较长键长。顶部(赭色):代表通过H2/Fe2+相互作用的理论进一步延长。左侧的箭头展示了Fe辅基从卟啉环的距离增加/减少。
结构表征进一步确定了高自旋和低自旋Fe-卟啉衍生物,研究表明,铁(Fe3+)也可以在两种自旋态中找到(Nihei等人,2007)。当发现高自旋Fe3+的键长增加了0.4 - 0.5Å,起始键长为2.04Å,远离卟啉的氮部分,与低自旋Fe3+的键长1.99Å相比(Rovira, Ballone和Parrinello, 1997; Bren等人,2015),这些实验得到了进一步扩展。因此,关于氢气相互作用是否有利于高自旋态Fe+2/3的问题产生了。
氢气是通过延长Fe+2/3键的长度,优化O2吸附通过稳定蛋白质结构,通过直接与Fe2+/3+辅基部分的相互作用,还是与人血红蛋白中的组氨酸93(以及其他蛋白质中相应的部分)的相互作用来发挥其效果,这一点尚未阐明。氢气也可能阻碍O2从Fe2+解离,从而改善O2饱和度,正如Singh等人(2021)所报道的,这一主题在第4.3.3节中考虑。
4.3.3 蛋白质口袋和空腔
关于氢气在躯体分布和/或作用的另一个主流理论是蛋白质口袋。这一假设基于在蛋白质结构内识别出离散的疏水通道和表面口袋。这些特征通常由亮氨酸、异亮氨酸、丙氨酸或缬氨酸等氨基酸排列而成,典型地形成于长度超过100个氨基酸的蛋白质中(Roose, Zemerov和Dmochowski, 2018)。疏水口袋能够与氩(Ar)、氪(Kr)、更大的原子氙(Xe)(Prangé等人,1998; Hancock等人,2022)以及可能是分子氢进行非共价/范德华相互作用。
与氢气类似,惰性气体通常被视为生物系统中的惰性物质,因为它们具有填满的电子轨道,意味着它们不参与经典的电子交换。然而,许多研究表明,Ar(Ye, Zhang和Sun, 2013; Nespoli等人,2019)、Kr(Perov等人,2021)、Xe(Lawrence等人,1946; Maze和Laitio, 2020)和氦(He)(Sykes和Lawrence, 1938; Manuilov等人,2022)可以具有神经和心脏保护效果。尽管,这些效果是如何引发的还没有完全阐明。下面的段落探讨了间接或非经典蛋白质/气体相互作用的可能性。
使用X射线晶体学评估氙-蛋白质相互作用的研究报告显示,疏水性和输送气体的体积是决定气体-蛋白质结合的主要因素,通过弱伦敦分散力发生(Abraini等人,2014)。伦敦分散力是一种弱量子力,涉及通常电性对称的原子之间的临时分子间吸引,导致非极性分子中形成暂时偶极(Lu, Neese和Bistoni, 2018)。因此,伦敦分散力可能部分解释了惰性气体的生物学模式。这种推测纯粹是学术性的,如果这种猜测要被反驳或支持,将需要进行广泛的建模和分析。
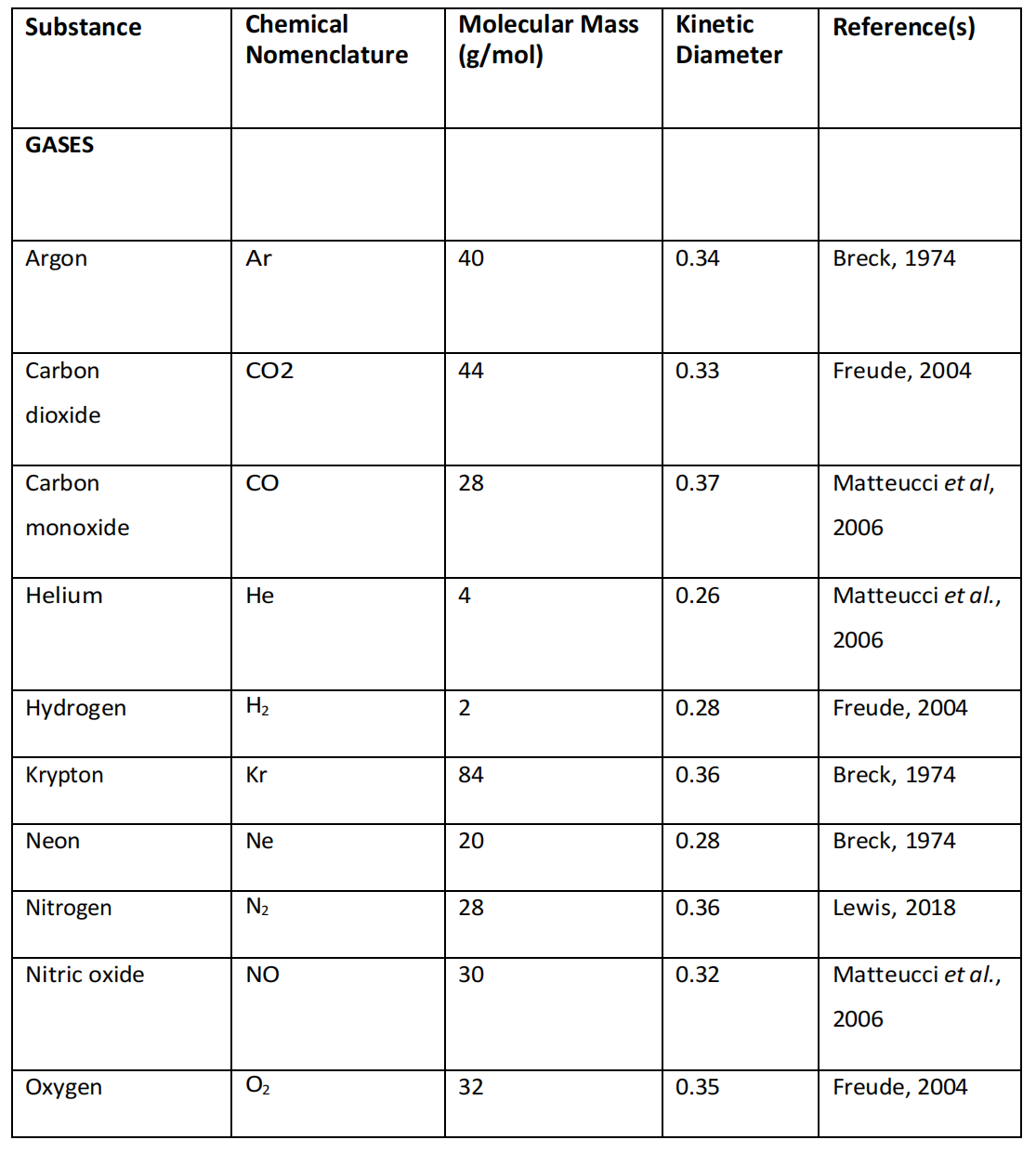
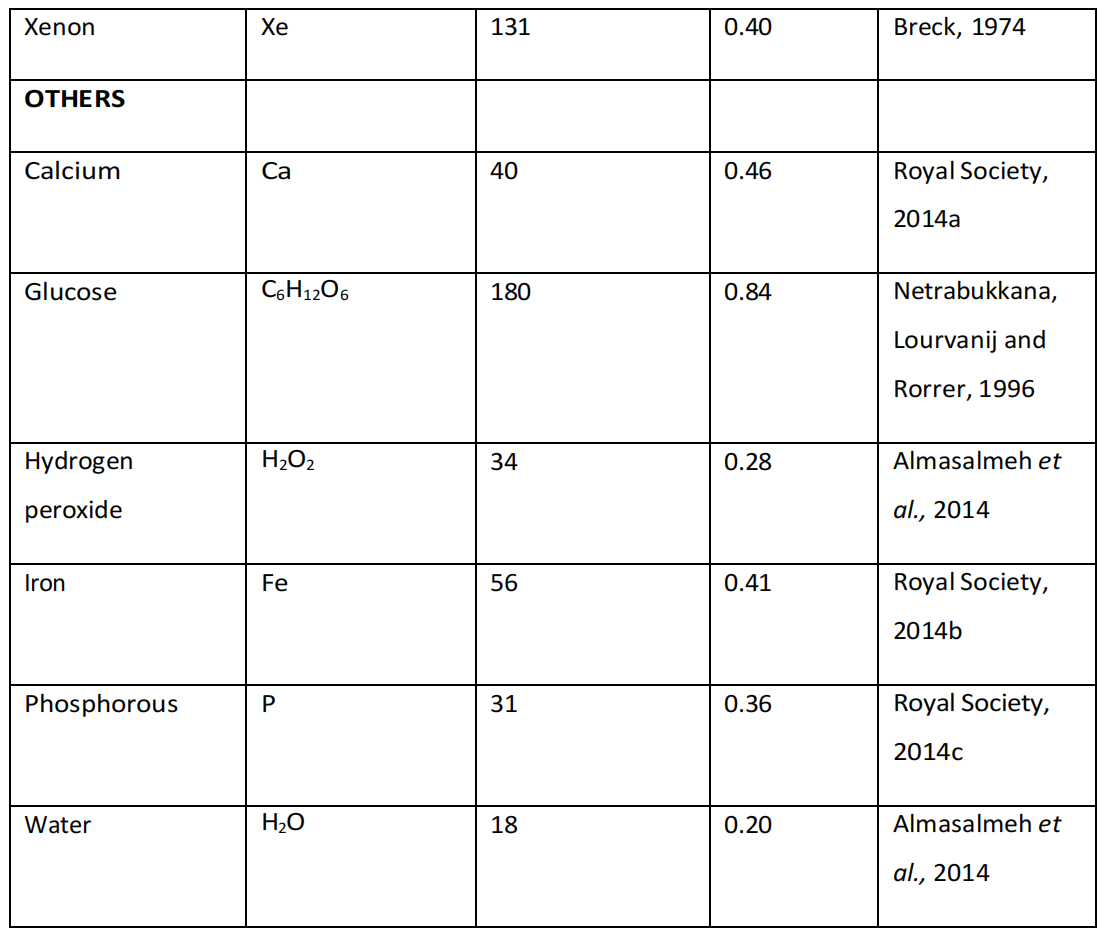
在评估蛋白质/分子相互作用的非化学效应时,应考虑的其他因素包括原子和分子的大小和重量,因为这些可能会决定这些分子影响蛋白质形态和功能的能力(表4.4)。
表4.4. 相关生物物质的质量与直径,注明生物分子的物理性质、原子质量及动力学直径(非水合、非离子态)。
研究鲸精液中提取的含有血红素的肌红蛋白的氧化形式——高铁肌红蛋白,观察到蛋白质内有四个独立的氙(129Xe)结合位点(Tilton, Kuntz和Petsko, 1984)。每个氙结合的蛋白质内通道都位于与装饰性铁相邻的氧气结合位点的对侧。此外,后来的报告识别出两个通道,通过这些通道氧气和氙都能穿过蛋白质到达它们各自的结合位点,这在结核杆菌的血红蛋白中已被确认(Cazade和Meuwly, 2012)。二氧动态的模拟将通道1识别为这些分子和一氧化氮的进入通道,而通道2主要被用作出口通道,尽管这个特定的通道被认为是双向的。作者还证明,氧气在穿过空腔到达血红素界面、对接位点(DS2/活性位点)时利用了氙口袋Xe1a和Xe2。有趣的是,一氧化氮只占据了Xe2位点,这可能通过抑制氧气通过通道2释放而有进一步的影响(图4.4)。因此,氢气可能利用这样的空腔、通道和/或表面口袋来支持蛋白质构型,并扩展蛋白质/金属假体键(第4.3.2节),这些效应可能促进含血红素蛋白中的氧气结合,并保护或维持金属酶(如CAT,SOD)的催化功能。
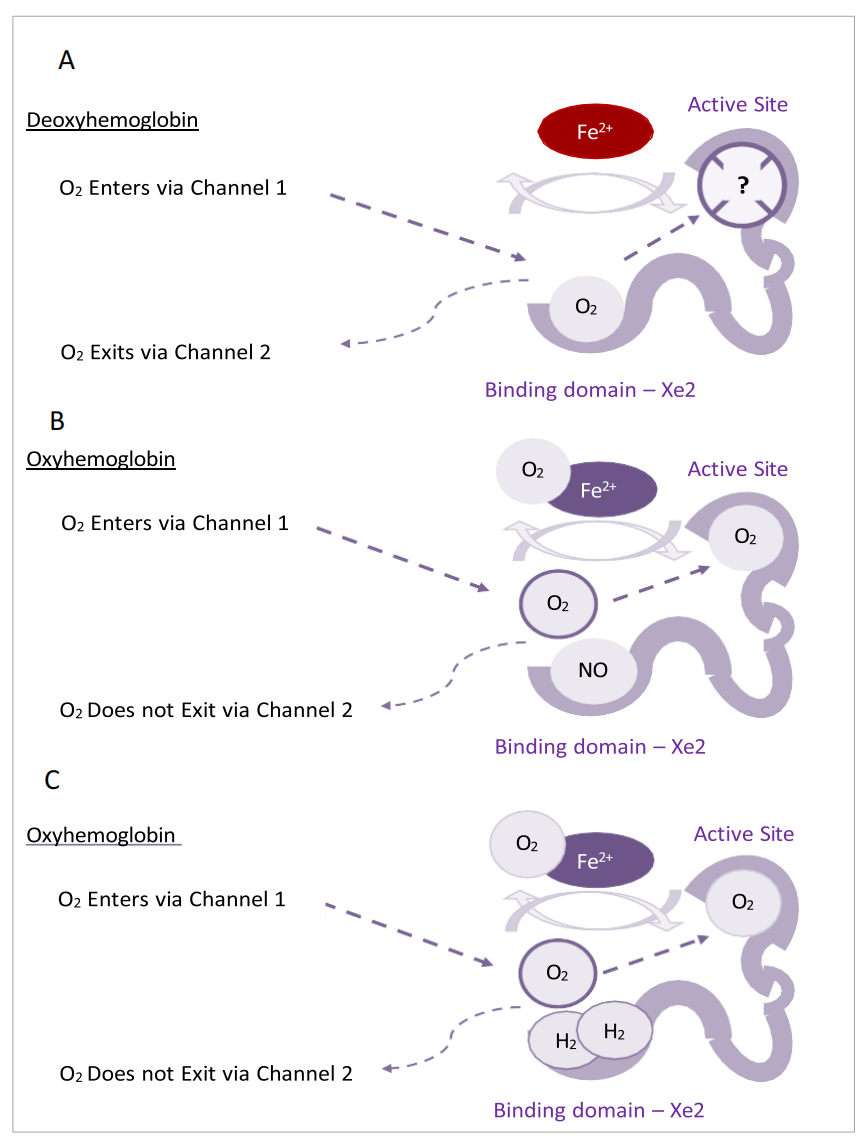
图4.4. 氙口袋“占用”对血红素氧合状态的可能相关性示意图。展示了分子如何与氙口袋相互作用以及可能的下游效应。活性位点:DS2对接域(Cazade和Meuwley, 2012)。(A). 脱氧血红素,氧气在被排出到出口通道前利用Xe2口袋。(B). NO在Xe2的结合阻止了氧气的结合,从而促进了血红素的氧合。(C). 与NO相似,氢气可能占据Xe2,导致血红素具有氧合潜力。
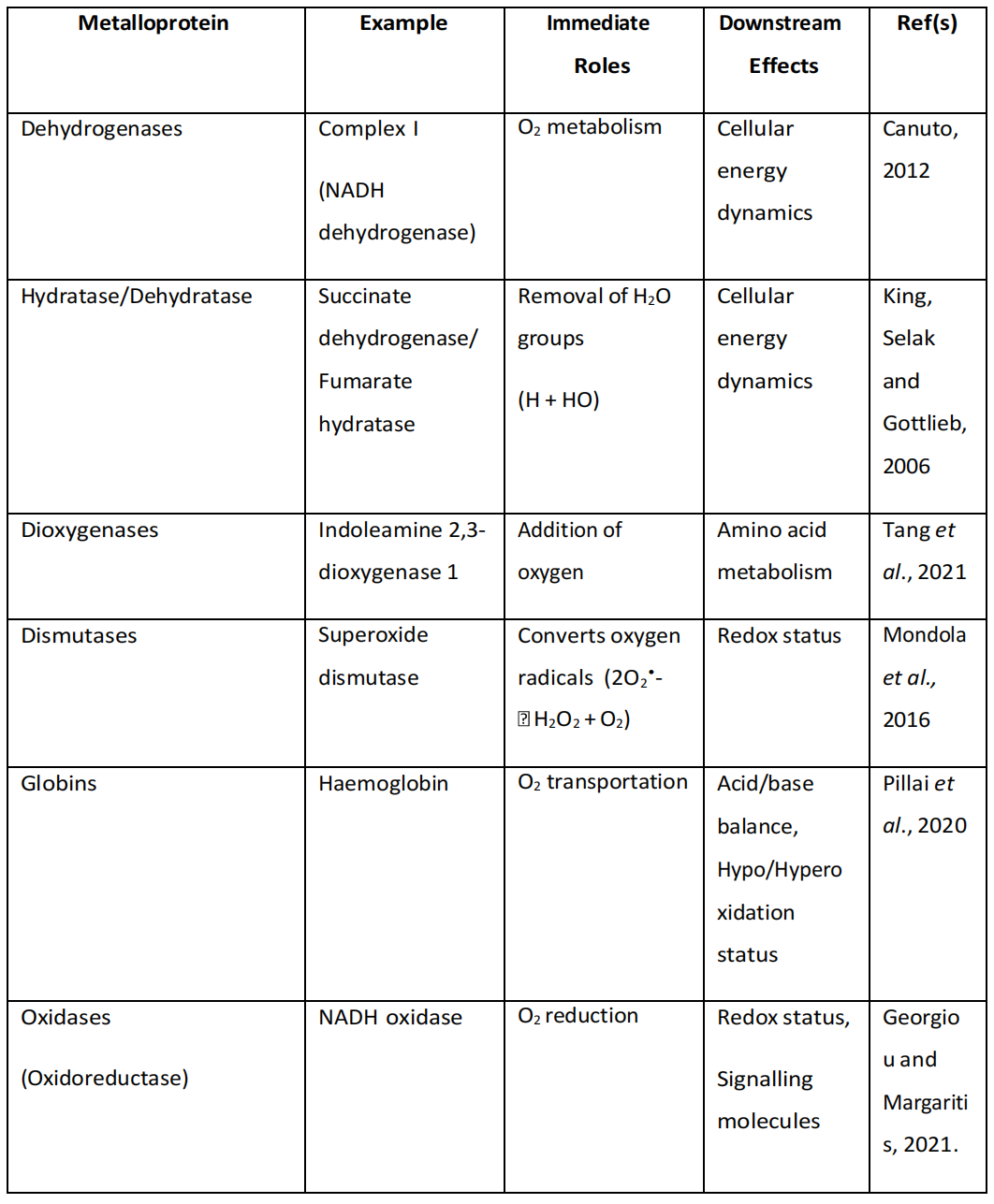
如果这样的过程确实在生理上被整合,那么可以推断在其他蛋白质单元中可能发生类似的过程。因此,如果铁磁性在氢气/蛋白质相互作用中扮演重要角色,那么下游效应也可能是全面的(表4.5)。
表4.5. 可能受氢气影响的金属蛋白的下游效应,确定了可能受氢气影响的金属蛋白的直接作用和下游效应。
支持氢气可能被吸引到铁磁性部分,并且至少暂时存在于蛋白质口袋中的观点,值得注意的是,出于工业储存目的,对氢气保留材料的研究往往集中在氢气分子的结构吸附上。在燃料电池中,碳纳米管(Hirscher和Becher, 2003)、富勒烯纳米笼(Shi等人,2020)和钯纳米晶格颗粒(Griessen, Strohfeldt和Giessen, 2016; Narayan等人,2017)因其优越且可逆的氢气保留性能而常被使用。含有金属离子的有机晶体结构被注意到具有高容量的氢气存储能力。金属有机框架是通过生物相容性金属离子(例如,Ca2+、Cu2+、Mn2+、Zn2+)与有机配体连接形成的微孔结构,能够在1 atm和1960C下保留相对大量的氢气,高达5.2% w/w(Rosi等人,2003;Suh等人,2012)。
使用这类微孔复合材料从生物学立场来看是有趣的,因为其结构通常提供与折叠蛋白质结构相似的微口袋。因此,根据氙口袋理论关于氢气活性和分布的说法(Hancock等人,2022;Hancock, 2023),氢气可以存在于这样的空腔中,或许与金属元素有相互作用。此外,如果这种现象发生在血红蛋白以外的蛋白质中(例如,参与生物分子合成、DNA修复和复制以及/或代谢和能量生产的蛋白质)(表4.5),它可以解释氢气的多效性效应,并说明氢气在组织内的移动。也许,氢气活性的蛋白质口袋机制可能具有一定的有效性。因此,关注含血红素蛋白与氢气相互作用的经验数据,通过核磁共振等技术以及计算模拟收集的数据将是非常宝贵的,如果能够解答以下问题:i)金属蛋白可能是氢气生物学效应的主要目标吗?ii)通过金属蛋白吸收在370C的生理温度下是否实现了生物学显著水平的氢气保留?
4.3.4 葡萄糖和糖原
葡萄糖是一种六碳单糖,化学组成为C6H12O6,在动物、细菌、真菌、植物和原生动物中普遍用作能源。葡萄糖是这些生物体中进行有氧和无氧代谢的关键底物(Hantzidiamantis和Lappin, 2022)。另一方面,糖原是一种由众多通过两种糖苷键连接的葡萄糖单元组成的大型支链多糖存储复合物,i) α-1,4-糖苷键和ii) α-1,6-糖苷键。在植物中,一种类似的结构称为淀粉,提供碳水化合物的存储能力。与糖原的支链结构不同,淀粉由两种聚合物组成,i) 形成线性和卷曲链的直链淀粉和ii) 形成支链的支链淀粉(Wang和White, 1994)。尽管糖原和淀粉的结构不同,但它们在化学上是相同的,都在各自的生物体中充当能量储存(Brust, Orzechowski和Fettke, 2020)。也许,由于这种化学相似性,糖原和淀粉都可能具有类似的氢气保留能力。
除了蛋白质结构(第4.3.3节),糖原复合物已被确定为有效的氢气保留分子,这在啮齿动物的肝脏和骨骼肌中得到了证明(Liu等人,2014;Yamamoto等人,2019)。肝脏是糖原产生的主要来源,这一特定器官中的氢气浓度(29μmol/L)被注意到超过其他系统,包括大脑(26 μmol/L)和肠系膜(23 μmol/L)(Yamamoto等人,2019)(表1.1)。有趣的是,已发现分支的糖原复合物是有效的氢气保留分子(第4章,第4.3.4节),这可能解释了肝脏中延长的活动(Liu等人,2014)。例如,对啮齿动物模型进行的3% 氢气吸入研究表明,心脏和大脑是氢气的近端和主要目标,但在肝脏和肠系膜中也发现了升高的氢气水平(Yamamoto等人,2019)。消耗氢富水(HRW)被证明可以减少耐力运动实验室模型中肝脏的糖原利用(Kawamura等人,2019),这表明氢气可能抑制糖原分解或促进葡萄糖转运进入细胞,从而抵消了葡萄糖释放的需求。
在如血浆或血清的水溶液中,由于高含量的氢键,葡萄糖分子变得粘稠(即粘性)(Te, Tan和Ichiye, 2010),因此氢气可能暂时与糖原或糖蛋白的碳水化合物部分结合(图4.5),也许也在红细胞表面(图4.6)。可以假设,可以与糖原的葡萄糖组分(或蛋白质的葡萄糖单元)形成弱非共价附着,将氢气分子困在其结构中,这可以解释远端部位以及肝脏和骨骼肌系统中氢气水平升高的原因,这些系统是糖原的主要储存处。此外,如果胶化是氢气可以粘附(也许是暂时地)到碳水化合物分子的一种机制,那么可以合理假设,在植物中与淀粉相同的效果也会看到(图4.5)。
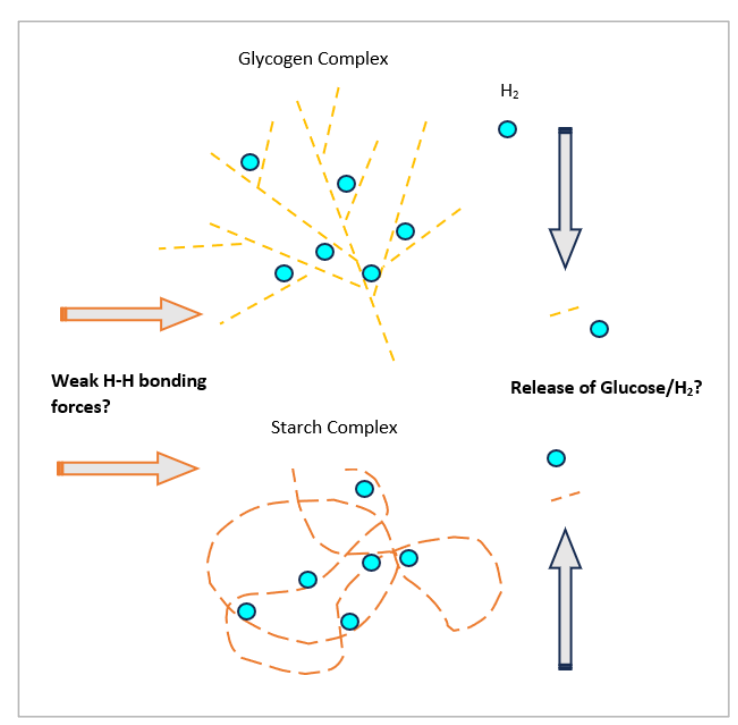
图4.5. 糖原和淀粉复合物中氢气保留和释放的理论机制展示了一个示意图,展示了氢气如何可能被糖原和淀粉存储分子捕获和释放。上图。糖原(动物、细菌、真菌)。下图。淀粉(植物)。
如果葡萄糖单元可以保留氢气,那么也可以合理假设氢气可以与游离葡萄糖或糖蛋白(例如,红细胞表面的血型糖蛋白A-D)结合,也许防止糖基化或其他翻译后修饰。考虑到解离,这种微弱的化学相互作用可能会被血流的速度冲刷掉(图4.6),在远端器官附近释放氢气。然而,这种推测需要经过实证评估才能验证上述理论是否成立。
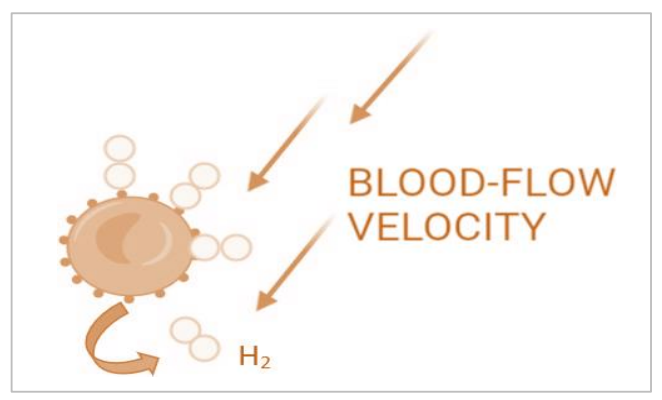
图4.6展示了一个理论化的氢气分布机制,通过示意图说明了氢气可能如何被红细胞上的糖脂/蛋白质的甘油基团捕获和释放。
4.4 细胞内信号传递
4.4.1 钙信号传递
氢气是一种非极性的二原子分子,这一因素降低了其在生物流体中的溶解度(Roose, Zemerov和Dmochowski, 2018),但增强了分子穿越生物膜并影响细胞内过程如信号转导(Itoh等人,2011;Begum等人,2020)、基因表达(Morgan等人,1997;Ohta, 2023)的能力,或许还有蛋白质活性和/或上述讨论的构象。在考虑细胞信号传递事件时,可以说负责调节细胞活动的最突出的两个信使是带正电的钙离子(Ca2+)和带负电的磷酸根离子(PO3 2-)(Clapham, 2007;Prakriya, 2020)。Ca2+调控一系列广泛的细胞事件,包括细胞收缩性和运动性、能量代谢和神经元活动,而磷酸根离子负责蛋白质的可逆翻译后修饰(由激酶催化添加磷酸根离子和由磷酸酶催化移除磷酸根离子),这一作用已知会改变蛋白质的极性,从疏水性(非极性)变为亲水性(极性),从而促进蛋白质-蛋白质相互作用(Prakriya, 2020)。钙信号传递对于最佳细胞功能至关重要,可以在细胞质、细胞器和细胞外环境中产生广泛的影响。钙信号传递是许多细胞类型中规范应激反应的基础,包括依赖Ca2+流入的内皮细胞和上皮细胞直接和间接地通过膜结合离子通道(Filippini, D’Amore and D’Alessio, 2019)。细胞对钙信号传递的反应在很大程度上取决于Ca2+的浓度、空间定位和化学物理特性。Ca2+分子高度移动,以40 nm/ms的速度在盐水溶液中迅速移动(爱因斯坦,1905年)。因此,严格调节钙信号传递以确保高度空间限制的信号传递,细胞使用一系列蛋白质反向转运蛋白、离子通道和泵控制局部浓度来螯合、分隔或排出Ca2+(Clapham, 2007;Thor, 2019),其中任何一个或全部都可能受到氢气的影响。
有趣的是,工业上生产保留氢气的纳米颗粒可以涉及用一层Ca2+涂覆预先形成的结构,以增强和加固氢气吸附(Yoon等人,2008;Beheshti, Nojeh和Servati, 2011),表明氢气可能直接与这些离子相互作用。从头计算表明,氢气可以通过弱电子供体力与氧化钙(CaO)的Ca2+组分结合,其中电子从占据的氢气σ轨道转移到未占据的Ca2+3δ轨道(Kim, Sun和Zhang, 2009)。如果这种结合在生理条件下发生,那么可以假设Ca2+和氢气之间存在强烈的亲和力,氢气可能会影响Ca2+-钙调蛋白(CaM)的结合潜力,这将对基因转录、免疫反应和肌肉收缩产生重大下游效应(图4.7)。钙调蛋白(CaM)是一个高度保守的Ca2+传感器,在细胞信号传递中具有基础性作用(Hussey, Limpitikul和Dick, 2023)。双叶结构的CaM分子可以结合多达四个Ca2+分子,每个结合区域对蛋白质构象产生个体效应,从而影响进一步的Ca2+结合和目标识别(Kawasaki, Soma和Kretsinger, 2019)。
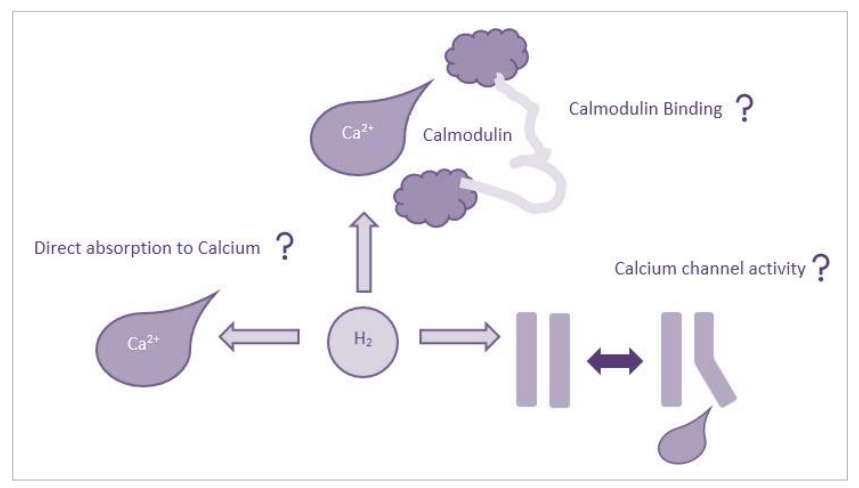
图4.7. 氢气给药对Ca2+信号传导的潜在影响。示意图展示了氢气如何影响Ca2+信号传导
尽管关于氢气与Ca2+及其衍生的信号级联反应的相互作用证据很少,但Iuchi等人(2016)证明,在纯化学系统中,氢气可以防止不饱和脂肪酸的自氧化,这一因素减少了Ca2+信号传导和下游Ca2+调节基因表达。该团队推测,对Ca2+信号的影响可能是由于激动剂诱导物或拮抗剂抑制剂的减少所致,尽管并未推测氢气是否可以直接与钙离子相互作用(Iuchi等人,2016)。氢气/Ca2+的直接相互作用也可能影响ROS的产生和活性,因为Ca2+过载会对克雷布斯循环酶的功能产生负面影响,激发ROS生成酶的活化(例如,α-酮戊二酸脱氢酶);并通过间接激活一氧化氮合酶(NOS),一个可以有效抑制细胞色素c氧化酶(复合体IV)功能、增加电子泄漏并最终促进线粒体功能障碍的因素(Feno等人,2019)。
或者,氢气可能通过上游效应间接影响钙信号。例如,氢气可能影响离子通道功能。如果是这样,氢气活动的特征是否类似于金属蛋白和酶中讨论的机制?例如,利用蛋白质口袋(第4.3.3节)的氙气被认为可以抑制钙信号传导(McGuigan等人,2023),因此氢气能否利用相似或不同的机制来调节Ca2+信号传导?
为了阐明这个难题,Itoh等人(2011)提出,氢气治疗抑制了LPS诱导的凋亡信号调节激酶1(ASK1)及其下游效应器蛋白(如c-Jun N端激酶(JNKs)、p38)在巨噬细胞中的磷酸化,而不影响到NOX-1产生的ROS。因此,氢气相互作用的潜在靶点可能位于受体蛋白或其下游。这些发现是引人入胜的,因为它们表明i) Ca2+直接或间接地可能成为氢气的目标,细胞水平直接受到跨膜离子通道和反向转运体配置的影响,以及/或Ca2+的大小、形状和电性状态;ii) 氢气作用的主要机制可能与氢气的直接还原潜力无关,而是通过调节Ca2+流入或活性。因此,应该考虑“氢气是否通过调节Ca2+流入来抑制ROS/RNS生成?”的问题。
4.4.2 蛋白质磷酸化
蛋白质磷酸化是真核细胞内信息传递的一种主要机制。磷酸化涉及通过添加或移除磷酸根离子磷酸基团对氨基酸(丝氨酸、苏氨酸、酪氨酸或组氨酸)进行可逆的翻译后修饰(Newcombe等人,2022)。将磷酸根离子分子添加到蛋白质残基上会改变被修饰蛋白的构象结构,从而改变其功能。激酶酶负责促进目标氨基酸的添加,而磷酸酶则催化分子的移除。
激酶已知形成级联,其中前体蛋白的磷酸化导致下游蛋白的连续磷酸化。MAPK/ERK级联是一个很好的例子,它们在细胞内外响应多种刺激(包括细胞因子、生长因子、有丝分裂物质如病原体相关化合物(例如LPS)和氧化应激)之间进行介导(Son等人,2011)。MAPK/ERK是一组与信号转导相关的超群蛋白的术语,这些蛋白在真核生物种类中高度保守。MAPK通路利用许多特征明显不同的蛋白质(例如,ERK 1/2、JNK、p38)通过细胞传达信号至可诱导的(例如,Nrf2)或转录元素(例如,ARE),这进一步影响细胞活动。例如,通过细胞外刺激或GTP结合蛋白激活的丝氨酸/苏氨酸MAPK3诱导MAPK2的磷酸化和随后的激活。然后,MAPK2参与MAPK激活环内的保守Thr-X-Tyr基序的双重磷酸化(Cargnello和Roux, 2011)。MAPKs在氨基酸残基上磷酸化目标底物信号转导,这是考虑超过13000个人体蛋白通过磷酸化/去磷酸化事件改变时的一个重要因素(Vlastaridis等人,2017)。促氧化细胞环境已知启动MAPK通路信号传导,通过激活细胞分化、运动性、增殖和生存途径,驱动细胞朝向保护和保存反应(Cargnello和Roux, 2011)。
因此,如果氢气能够通过累积在蛋白质口袋中(第4.3.3节)或直接作用于磷酸基团或磷酸化的目标(例如,丝氨酸、苏氨酸等)来抑制磷酸化级联,也许通过量子力(例如,伦敦分散),它可以为当代文献中注意到的多效性效应提供一个统一的理论(Itoh等人,2009;Xia等人,2013;Zhuang等人,2020;Slezak等人,2021;Li等人,2022)。特别是,p38是一种与炎症反应增强有关的进化上高度保守的MAPK蛋白。例如,p38活性增加与更高的恶性风险有关,而MAPK/p38抑制剂被证明可以减少炎症,这是肿瘤疾病的标志(Naser等人,2023)。氢气被证实可以在体外利用体细胞培养(Begum等人,2020)和配子(Ma等人,2022),以及在体内,使用啮齿动物疾病模型(Liang等人,2012;Wang等人,2020b)抑制MAPK/ERK/JNK/p38信号传导。尽管观察到的氢气应用效果是由于磷酸酶活性增加、激酶活性减少、NFκB/TNF-α活性降低还是另一种尚未定义的机制(图4.8),这一点尚不清楚。
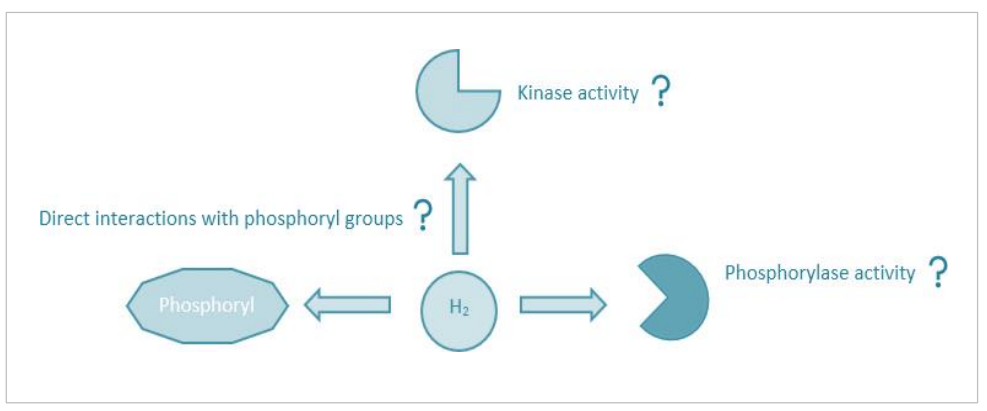
图4.8. 氢气给药对磷酸根离子信号传导的潜在影响。示意图展示了氢气如何影响磷酸根离子信号传导
4.4.3 磁电化学
生物磁性研究,即生物体产生的磁场,已经非常成熟,心脏和大脑的磁场、心磁图和脑磁图分别在临床实践中被常规使用(Roth, 2023)。
众所周知,磁力可以改变电子自旋的动力学。
在此,讨论了铁磁性辅基团的量子力,如血红素(Fe)或SOD(Mg, Cu),是否影响自旋态,从而影响氢气的生物学活性的问题。
已知氢气的热极化有利于三重态电子状态,大多数氢气分子在室温下被观察到处于正交态,或三重态(约75%正交/25%顺磁自旋)(O’Neill等人,2023)。由于正交电子自旋产生弱抗磁性,排斥力(Tarzad, 2020),它不太可能直接减少ROS,因为一种力可能会排斥另一种力。然而,三重态自旋氢气分子可能影响细胞生物化学的某些方面,如基因表达、蛋白质构象和/或信号转导。尽管目前是一种推测性假设,但对驱动氢气生物学相互作用的量子力的深入分析可能提供有价值的数据。
虽然大多数生理氢气可能具有抗磁性,但顺磁态或单态氢气的影响同样重要。占生理氢气约25%的顺磁自旋氢气分子产生弱顺磁性,或吸引力。单态可能通过磁电化学吸引金属蛋白和辅基团而具有氧化还原活性潜力。当正向电子转移被抑制时(例如,线粒体复合物I),会发生自由基形成,这有助于ROS的形成(Hore, 2012)。例如,一个黄素-超氧自由基对(FH• + O2 •- )带有单态自旋(O2),将形成F + H2O2,而同样的对在三重态自旋状态下会解离,导致两个自由基分子,FH•和O2 •-(Ramsey和Kattnig, 2022)。因此,如果氢气的电子处于单态配置直接与蛋白质的过渡金属组分相互作用,正如Hancock和Hancock(2018)、Kim等人(2022)和Ohta(2023)所描述的,那么金属组可能通过降低游离氢气的解离能(约4.64 eV - 约2.35 eV)和形成受体/H•复合物来催化自由基的还原。逻辑上,H•自由基可以与氧化剂如O2 •- 和•OH反应,尽管正如第4.2节所述,这将取决于所感知到的自由基的空间和时间可用性。关于氢气的磁电化学效应进一步考虑的因素包括CAT和SOD等抗氧化酶的金属催化中心,据报道在暴露于氢气后活性增强。可以推测,单态自旋氢气也可能通过提供额外的H•并保留催化金属元素的还原能力来改善这些酶的功能,尽管这还有待经验验证。
因此,由于微弱的磁力作用,单态分子可能具有抗氧化效果,而三重态氢气分子可能通过稳定蛋白质结构来影响诸如细胞信号传导、基因调控和代谢活动等细胞活动(第4.3.3节)。当推断三重态自旋状态时,氢气还可能通过促进或抑制蛋白质磷酸化/去磷酸化事件来介导细胞信号传导,这很可能取决于蛋白质的原始功能和/或构型。如果这些理论概念是准确的,它们可以解释氢气在活体系统中的多效性效应。而且,正如Ohsawa等人(2007)、Barancik等人(2020)、Tian等人(2021)和Jin等人(2023)所建议的直接抗氧化活性,以及Itoh等人(2007)、Wilson等人(2017)、Itoh等人(2019)和Slezak等人(2021)描述的对细胞信号传导和基因调控的影响,都可以得到调和。
https://wap.sciencenet.cn/blog-41174-1450502.html
上一篇:免疫系统的性别差异
下一篇:大爆炸过后,如何燃烧掉无处不在的氢气?