博文
我们不知道答案的125个科学问题(105)植物免疫的适应性  精选
精选
||
105. 为什么植物不能对所有的疾病都免疫?
Why aren’t all plants immune to all diseases?
题记:植物和动物不同它只具有一般的先天免疫反应,不存在后天的适应性免疫系统,但植物也可以通过变通的分子抗体蛋白来识别和消灭特定的病原体。植物病理学家不明白为什么不同的植物物种,即使是近亲,也会有不同的防御系统。该问题的答案可能会让人类种植出更加强壮抗病的农作物。
植物和动物一样会生老病死,但由于植物不能移动而主动地去逃避病虫害的伤害,所以植物发展出强大而复杂的先天免疫系统,其免疫功能主要依赖于每个细胞的先天免疫和从感染部位发出的系统信号来运作。由于植物强大的先天免疫系统,其对周围环境中的大多数疾病都有强大而持久的免疫力,这种神秘的特性被称为植物的非宿主抵抗力(non-host resistance, NHR),正因为这种NHR的存在,植物能有效地抵御真菌、细菌和其他聚集在空气、土壤和身体中各种病原体的不断攻击,在大多数情况下,植物对这些病原体都能成功免疫,而植物成功激活NHR是因为它能对DNA酶产生反应(DNA酶是一系列催化DNA分子变化的酶的总称),而大多数病原体线粒体中的DNA酶都有一个小肽分子,它们能够顺利穿过植物细胞膜,从而诱导植物NHR的激活表达。

图1 植物的真菌叶斑病
尽管植物具有坚硬复杂的细胞壁和强大的先天免疫体系,但它依然不能对所有的病原体都能产生有效的免疫。植物是生物界最重要的营养物质的来源,所以与植物寄生相关的微生物是损害植物生长和繁殖的主要病原体,比如线虫和蚜虫通过将柱头直接插入植物细胞来取食,而真菌可以直接进入植物表皮细胞,可以在植物细胞上、细胞间或通过植物细胞延伸菌丝。病原真菌和共生真菌及卵菌可以内陷取食器进入宿主细胞质膜,其吸器质膜、细胞外基质和宿主质膜形成一个亲密的界面,在这个界面上真菌可以汲取营养不断发展。另外致病菌可以通过植物的气孔、水孔或伤口进入组织,它们在细胞间隙(外质体)增殖。这些不同种类的病原体都将效应分子或毒性因子(virulence effectors)传递到植物细胞中,干扰植物的正常生长和代谢以增强微生物的在植物体内的生存适应性。作为回应,植物因此进化出了抗体(resistance, R)蛋白,可以检测这些毒性因子所诱导的扰动信号,进化出通过少量R蛋白特异性识别大量具有相似感染策略的病原体的能力。
为了应对这些不同的病原体,植物进化出复杂的多种免疫反应体系,如模式触发免疫(PAMP-triggered immunity, PTI)、效应因子触发免疫(effector-triggered immunity, ETI)、RNA默化(RNA silencing)和自噬(autophagy)(参考: Plants. 13:1434, 2024),这些复杂的免疫反应由多种信号级联进行调节。植物中最常见的免疫系统反应主要是PTI和ETI,称为双支先天免疫系统(见图1所示):PTI分支通过细胞表面受体识别并响应许多种类微生物共同的分子模式,包括非病原体;ETI分支通过细胞内受体对病原体的病毒因子(virulence effectors)作出直接或间接反应。也就是植物主要的两种先天免疫通道根据受体激活模式可分为微生物模式识别和病原体效应蛋白的识别两大类,PTI模式是使用跨膜模式识别受体(pattern recognition receptors, PRRs)对缓慢进化的微生物或致病菌的相关分子模式(microbial- or pathogen-associated molecular patterns, MAMPs or PAMPs)做出识别反应,如鞭毛蛋白等;ETI模式则主要在细胞内发挥作用,主要使用大多数R基因编码的多态NB-LRR (nucleotide-binding, leucine-rich repeat, 核苷酸结合的、富含亮氨酸的重复体)蛋白产物对病原体进行“特异性识别”并激活防御反应(NB-LRR只针对宿主型病原体有效,如果病原体杀死宿主细胞如坏死性植物营养菌无效,参考文献: Nature 444:323–329, 2006)。大量的研究证明植物免疫的这两个分支并非是两个完全彼此独立运行的免疫通道,它们的协同免疫反应通过图2所示的方式进行。
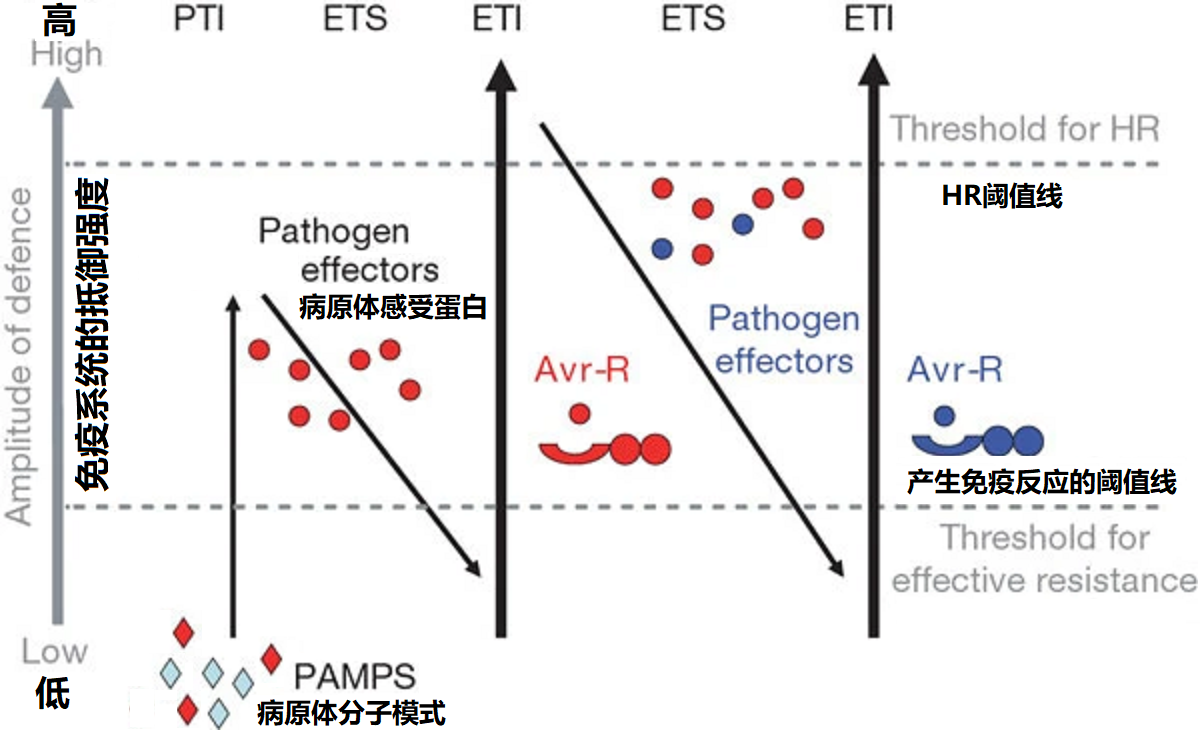
图2 植物的免疫反应过程
如图2所示,在植物免疫反应的第一阶段,植物通过PRRs检测微生物或致病菌相关的分子模式(用红色钻石表示),从而触发PTI免疫;在第2阶段,成功感染植物的病原体会产生干扰PTI的效应物(干扰素),致使病原体获取营养和进行扩散,造成病原体感受蛋白产生的易感性(effector-triggered susceptibility, ETS)阻断植物PTI响应,导致免疫防御能力降低;在第3阶段,病原体感受蛋白(红色圆)被NB-LRR蛋白识别,从而激活了效应因子触发免疫(ETI),这是一种比PTI更强烈的免疫反应,通常会突破超敏细胞死亡(hypersensitive cell death, HR)的阈值,导致感染部位的超敏细胞死亡。在第4阶段,被识别的病原体被隔离淘汰而大大减少,这样病原体为了生存发生了变异,从而产生了新的R特异性,躲避植物的ETI反应。作为回应植物体又通过获得等位基因(蓝色)产生新的R感受蛋白来识别这些变异的病原体,从而再次导致激活ETI,这种相互作用使得植物和病原体之间形成了复杂的相互选择和进化关系。
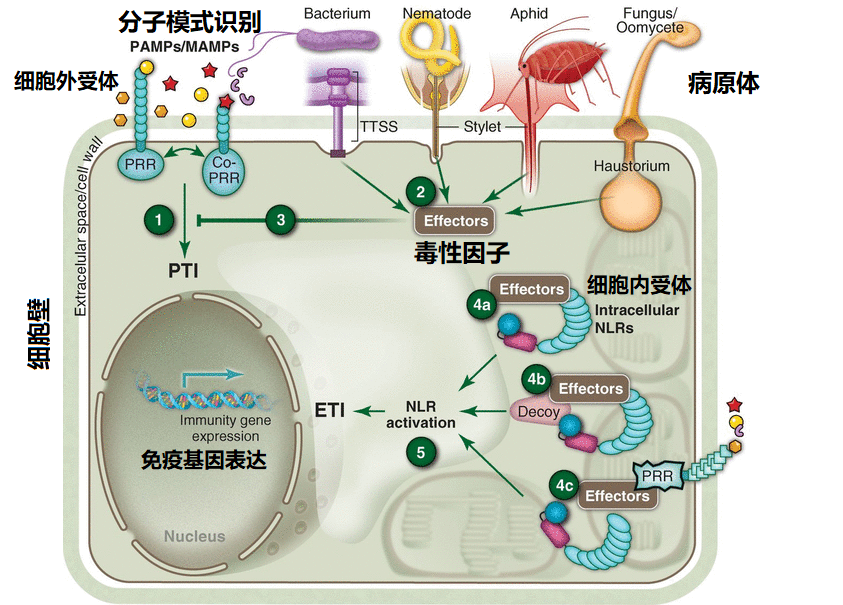
图3 植物免疫系统示意图
虽然植物体缺少后天的适应性免疫系统,但是植物可以启动特异性的、自我耐受的ETI免疫反应,并因此建立免疫记忆。在这种免疫机制中植物体是通过特异性的抗体(resistance, R)蛋白来识别特定的某类病原体的,当某种病原体将自己的毒性因子注入植物细胞中时,植物就能通过R蛋白快速识别病原体。植物的这种抗体免疫能力可以通过物种内和物种间的杂交来遗传,从而让植物先天就具备这种对特种病原体的自身免疫能力。当植物的组织被病原体感染后,其组织细胞会产生多种脂类或激素类的信号分子,这些信号分支可以被运送到全身各个组织,从而让植物体整体获得全身免疫力,这个过程将涉及植物细胞中数千个基因的重编和调节,让植物在疾病感染后优先考虑免疫应答而不是正常细胞的常规需求。
虽然表观遗传修饰和位点特异性染色质重塑提供了植物对病原体攻击的持久记忆,而且这种病原体诱发的特定位点的基因组重排可以传递给后代。然而根据图2所示的植物免疫过程来说,植物通过ETI与病原体斗争的过程中,由于病原体与植物免疫系统的选择竞争,病原体多种多样而且迅速的演变会引起植物不同的疾病和流行病;所以病原菌分布和寄主免疫敏感性的差异,造成植物对病原体感染的反应存在很大的异质性,病原体在植物上的快速适应性使得不同物种甚至同近亲间的免疫防御系统也能产生很大区别,再加上植物缺乏循环系统和可移动的免疫细胞,它们就不能使用循环免疫受体来检测和追踪特定的细菌和病毒,造成植物不同感染部位的R蛋白都能产生差异。尽管如此,植物依然能够建立高度特异性的免疫反应,也具有有限的自我反应性,并且通常会对遇到的病原体产生终生的“记忆”。所以植物与病原物之间长期存在的复杂的相互选择和适应, 使得病原体的致病性和植物的抗病性都呈现多样性,这就决定了为什么不同的植物物种,甚至相关密切的近亲物种对病原体具有完全不同的抗病反应。
* 植物免疫方面的细节问题非常多,具体可以参考2021年Nature期刊关于植物免疫领域的专题系列文章,共有七篇研究论文(中国2篇),该系列成果报道了在植物免疫和抗病研究中的一些比较重要的发现和结果。
https://wap.sciencenet.cn/blog-318012-1455948.html
上一篇:我们不知道答案的125个科学问题(104)植物生长控制
下一篇:我们不知道答案的125个科学问题(106)植物的抗逆性