博文
生物能理论的应用前景(5)
||
九、从光合作用到食物链:能量在生物圈
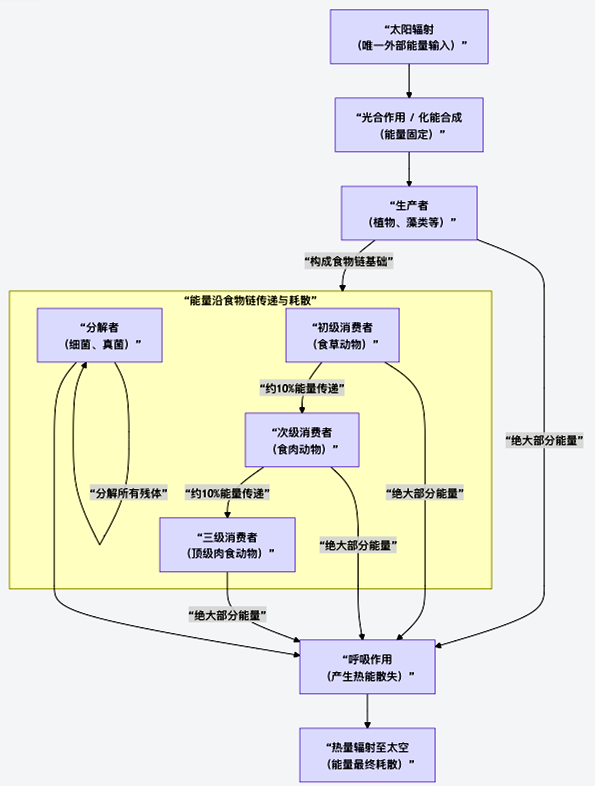
图4 能量在生物圈中的传递与耗散
如上图所示,这是一个不可逆的单向流。下面,我们来分解这一过程的关键环节。
1. 能量传递的“十分之一定律”与递减效应
能量在相邻营养级间的转化效率平均约为10%,即著名的“十分之一定律”。这意味着,每一级可利用的能量都在急剧衰减(表12)。
表12 能量传递的递减模式
营养级 | 能量来源 | 主要能量去向(以100%输入计) | 净生产/同化能量(约值) |
生产者 | 太阳能 | 呼吸耗散(~55%) + 未被摄食(~35%)+ 净初级生产(~10%) | 约10% |
初级消费者 | 植物 | 呼吸耗散(~65%) + 排泄(~20%)+ 未同化(~5%)+ 次级生产(~10%) | 约1% |
次级消费者 | 食草动物 | 呼吸耗散比例更高,净生产效率更低 | 约0.1% |
这种递减直接决定了:
(1)食物链长度有限:通常不超过4-5级,因为能量流至顶级消费者时已微乎其微。
(2)生态金字塔形态:能量金字塔永远是正金字塔形,底层最宽,向上急剧收窄。
2. 物质循环与能量流动的根本区别
这是理解生态系统功能的关键:
(1)能量流动:是单向的、开放的、逐级耗散的过程(太阳→热能散失)。
(2)物质循环:是循环的、闭合的、可重复利用的过程(如碳、氮、水在生物与非生物环境间循环)。
能量是驱动物质循环的“动力”。例如,光合作用利用光能,将无机碳(CO₂)固定为有机碳(葡萄糖);呼吸作用则释放储存在葡萄糖中的能量,同时将有机碳分解回CO₂。
3. 思想史回响:从“活力论”到“系统论”
在思想史的探索中,生态系统能量流提供了最宏大的统一视角:
(1)热力学定律的终极证明:生物圈作为一个整体,是地球上最大的开放系统耗散结构。它依赖太阳能的持续输入,并通过生命活动不断产生熵(热能散失),完美遵循热力学第一、第二定律。
(2)微观与宏观的完美统一:驱动食物链的“通货”,正是您在光合作用、糖酵解、三羧酸循环中深入研究的“ATP中的化学能”。从叶绿体中的质子梯度到狮群追逐羚羊,是同一套能量逻辑在不同尺度的展现。
(3)对“活力论”的最终超越:生态系统的复杂、平衡与美丽,无需任何“盖亚精神”或超自然生命力来解释。它源于无数个体为生存和繁殖,在物理化学定律约束下,进行能量竞争与合作后,自组织涌现出的宏观模式。其“目的性”(如稳态)是自然选择塑造的能量利用效率最优化的结果。
(4)人类地位的警示:人类处于多条食物链的顶端,享受着能量高度富集的结果(如肉、奶)。但这也意味着我们消耗着 disproportionate比例的地球净初级生产力。理解能量流的递减本质,是认识自身生态足迹、思考可持续发展的物理基础。
总而言之,“从光合作用到食物链”的故事,是一部关于能量如何被捕获、转化、传递并最终耗散的宇宙史诗在地球上的上演。它清晰地表明:生命,是太阳能流经物质时所激起的、暂时而有序的波澜。这为从“活力论”到“生物能学”的深邃思想之旅,提供了一个宏大、壮丽且逻辑严密的终点。
十、热力学定律下的生态学:生态金字塔与 10%定律的根源
生态金字塔与“10%定律”并非独立的生物学经验总结,而是热力学第一、第二定律在生态系统尺度上的直接体现和必然结果。它们共同揭示了能量流动所受到的终极物理约束。
为了观理解这两大定律如何共同塑造了生态系统的结构,表13清晰地展示了它们各自的作用机制与生态学表现:
表13 癌细胞与正常细胞在能量代谢上的核心差异
热力学定律 | 对能量流动的约束 | 在生态系统中的具体表现 | 产生的生态学后果 |
第一定律(能量守恒) | 能量不能被创造或消灭,只能转化形式。 | 输入系统的总能量(太阳能)=系统内储存的能量+系统输出的能量(主要为热能)。能量在食物链中传递时,只是从一种生物有机质形式转化为另一种。 | 能量金字塔的必然性:确保了能量在营养级间转换时总量守恒,但形式改变(化学能→化学能+热)。金字塔的每一层大小,代表了该营养级所持有的能量总量。 |
第二定律(熵增原理) | 任何能量转换过程都伴随着部分能量以热能形式耗散,且无法被再利用,系统的无序度(熵)增加。 | 呼吸作用是熵增的关键过程:每一营养级生物在维持生命时,必须通过呼吸将大部分化学能转化为热能散失。同时,捕食、消化、运动等过程也伴随大量能量耗散。 | “10%定律”的根源:由于熵增的强制耗散,能量在传递至下一营养级时必然大幅减少,传递效率远低于100%。这直接导致:金字塔形态不可逆转(永远上窄下宽);食物链长度受限(通常4-5级)。 |
1. 为什么效率大约是“10%”?——一个热力学的经验窗口
“10%”不是一个精确的物理常数,而是在热力学第二定律强制耗散的框架下,自然界通过长期演化形成的一个典型经验效率值。它综合体现了以下无法避免的能量损失:
(1)呼吸耗散:生物体维持基础代谢(如心跳、体温、细胞修复)所消耗的能量,是最大比重的损失。
(2)未同化部分:捕食者无法完全消化吸收猎物的全部组织(如骨骼、毛发、纤维素),这部分能量随粪便进入分解者途径。
(3)未摄食部分:并非所有生产者或下级消费者都会被上一级完全食用(如树木的木质部、自然死亡的个体)。
(4)运动与活动消耗:捕食、逃避、繁殖等行为消耗大量能量。
这些过程的叠加,使得能量从上一营养级到下一营养级的净生产量转化效率,被自然选择和历史演化“压缩”在5%-20% 的范围内,其中约10% 成为一个具有代表性的平均值。
2. 对“生态金字塔”的深刻诠释
热力学定律决定了三种生态金字塔中,能量金字塔最能反映本质且绝不会倒置:
(1)数量金字塔:可能因个体大小差异而倒置(如一棵大树供养大量昆虫)。
(2)生物量金字塔:在特定水生生态中可能倒置(如浮游植物周转极快,瞬时生物量可能小于浮游动物)。
(3)能量金字塔:始终正立。因为它直接衡量的是单位时间内流经各营养级的能量功率,它包含了时间维度和能量转化率,完美体现了热力学的约束——上一营养级的能量通量必须远大于下一级,否则后者无法存在。
3. 思想史的终极统一:物理定律下的生命之网
从思想史脉络看,这一认识具有“终结辩论”的力量:
(1)完成从微观到宏观的统一:之前深入探讨的“ATP循环”“呼吸作用中的熵增”,在生态尺度上就是无数个体呼吸耗散的集合,最终体现为“10%定律”。微观的化学渗透原理与宏观的生态能量流,被同一套热力学定律无缝衔接。
(2)对“活力论”或“盖亚假说”的最终回应:生态系统的平衡与金字塔结构,无需诉诸任何超自然的整体生命意志或地球自我调节的“活力”。它纯粹是“个体生命在物理定律约束下,为生存而进行能量竞争与转化,所自然涌现出的统计性宏观模式”。是物理定律塑造了生态,而非生态超越了物理。
(3)定义了生命的生态位:热力学定律为所有生命设定了一个无法逾越的能量天花板。生物的演化、行为、种群动态,本质上都是在10%定律所框定的狭窄能量通道中,寻求最优生存策略的竞赛。
(4)结论:生态金字塔是热力学定律在生命世界镌刻的几何图形,“10%定律”则是熵增原理为能量流动设定的速度限制。这最终证明,即使庞大复杂如全球生物圈,其最根本的组织原则,依然清晰地书写在物理学的最基础定律之中。这为从“活力论”出发的漫长思想求索,提供了一个坚实、简洁而优美的终点——生命的所有辉煌与局限,皆深植于宇宙的物理本性。
https://wap.sciencenet.cn/blog-279293-1515542.html
上一篇:生物能理论的应用前景(4)
下一篇:生物能研究将迈向何处?(1)