博文
A story between HBV and STING
 精选
精选
|
治疗乙肝的新靶点——STING
今天看到一则朋友圈的动态,转发了总理批的《“十三五”国家科技创新计划》,这次提及生物医药的次数较之前更多、更广、更新,重大传染病的防治依然列在前几项(记得“十一五”期间也是如此),这次主要突出了攻克两类疾病:乙肝和肺结核诊防治等关键技术和产品。看到医药魔方曾贴出的一则消息汇总了全球乙肝药物研发管线,一看便知,乙肝的药物研发确实如火如荼,后续将会有更多的团队参与角逐。
诱发乙肝的病原是乙型肝炎病毒(hepatitis B virus, HBV),属于嗜肝DNA病毒科(hepadnaviridae)中的DNA病毒,是最小的动物病毒之一,基因组长仅3.2 Kbp(比实验室常用的质粒还要小,却危害极大),为部分双链环状。HBV颗粒(也叫Dane particle)的结构比较简单:外层包膜上插有表面抗原(HBVsurface antigen, HBsAg),内部二十面体核衣壳蛋白组成的核心结构(Core),包裹着基因组DNA和DNA聚合酶(类似于反转录病毒的反转录酶,一种RNA依赖的DNA聚合酶。这也是病毒的神奇之一,是他们丰富了中心法则)。
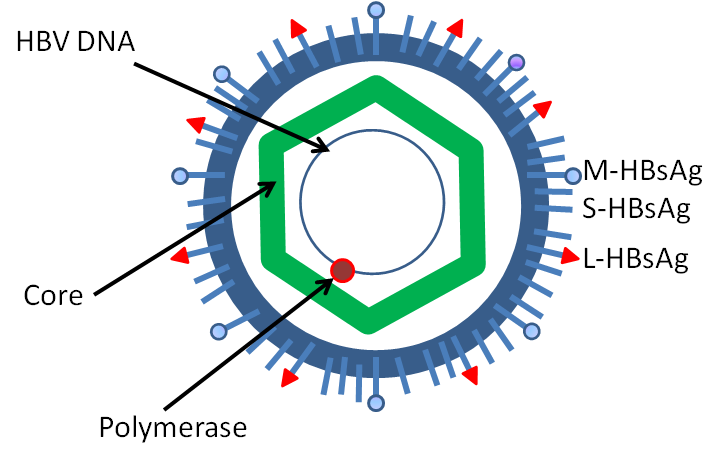
Simplified diagram of the structure of hepatitis B virus (by Dr Graham Beards)
曾经的研究方向属于分子病毒学,集中在反转录病毒科,曾关注过慢病毒属的HIV,也知道泡沫病毒(foamy virus, FV)的一些故事。可当时纯粹是为了做完手里的某个蛋白-蛋白的相互作用、或是某个DNA-蛋白的相互作用,却从未思考过要如何真正的为患者治疗这类疾病。可能是HIV过于强大,至今没有根治的方法,病毒和宿主的博弈愈演愈烈;可能是FV过于柔和,她可以感染诸多的灵长类和非灵长类的动物,却不致病,我们也就自然将目光集中在开发病毒载体这个领域或是研究她们不治病的机理。
现在来看,乙肝的疫苗具有90%的免疫保护效力,治疗却比较难,方法较为单一(多年来依赖于干扰素和核苷酸类似物)。国际上的药物研究已从多个靶点铺开,包括Gilead的核苷酸类RT抑制剂、TLR7激动剂和治疗性疫苗,Hepatera的病毒进入抑制剂、Replicor的HBsAg释放抑制剂、Arrowhead的RNAi疗法和靶向cccDNA的疗法、J&J的衣壳蛋白抑制剂、GSK的反义药物、还有Spring Bank的RIG-I和NOD2的激活剂等等,国内的研究还处于起步阶段,主要是干扰素类似物,临床试验上仅看到东阳光有开展衣壳蛋白抑制剂方向。其实还有一个应该是比较热门的靶点,目前还未被广泛报道——STING。
一个月前Invivogen的销售过来推销他们的新产品,主打的就是STING的系列细胞系和刺激物,也是那一次我才知道原来Invitrogen原来带有In vitro这个含义。记得那次报告从先天免疫(innateimmunity)的PAMPs和PRRs开始,到人们渐渐的揭示DNA sensor 和STING之间的递呈关系,再到后来人们发现细胞质中的DNA会被cGAS(cyclic GMP-AMP synthase)识别并转变成cGAMP(cyclic GMP-AMP),而这个小分子就是能激活STING通路的第二信使,接着向内部传递的信号:STING-TBK1-IRF3-IFNs。有IFN产生,自然和先天免疫密切相关。
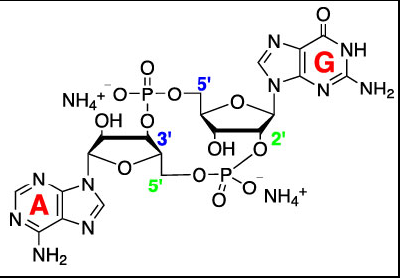
Metazoan CDNs, 2ˈ3ˈ-cGAMP MicrobialCDNs, 3ˈ3ˈ-cGAMP
今天突然很想知道HBV的DNA是否可以通过STING将信号传递下去?或者,上调STING的活性是否可以抑制HBV的复制,进而为其他药物的联合治疗提供备选方案?昨日的Friday Seminar中讲到OCT2这个转运蛋白活性上调后会提升癌细胞对药物的敏感性,可以明显看到体内模型中的肿瘤shrink。若是STING通过某种方式或分子得以上调活性,对乙肝的进程又是有如何影响呢?因为没有听人谈论过,瞬时觉得很有可能,赶紧Yahoo一下,看看别人都做了什么,目前已经到了何种程度。稍稍有些思绪,还是很想与大家分享。
HBV感染细胞后会产生cccDNA(covalently closed circular DNA,共价闭合环状DNA),他们是类似质粒的游离基因,来自与蛋白相连的、经反转录后产生的RC-DNA(relaxedcircular DNA),但如何从RC-DNA到cccDNA还不是很清楚。[1] cccDNA在细胞核中作为模板,指导HBV RNA的转录,也会被包装到病毒的颗粒中。HBV的持续性感染和cccDNA的存在,像是HBV最好的隐秘方式,潜伏于感染的细胞中,待时机成熟再大量复制、扩增。难以清除潜伏的病毒,也是AIDS治疗时让人头疼的地方。究其根本,主要是体内没有足够的免疫力清除这些外来的病原,自然就会让人们想到先天免疫和获得性免疫。有文章报道称HBV始终非常狡猾的病毒,他们有某种机制可以低水平的诱导机体免疫水平,逃避监控。从这个角度出发,如何增强机体的免疫能力,或许是一条抑制HBV复制、治疗乙肝的方向。[2]提升获得性免疫能力的方法有多种,比如疫苗激活(如重组的HBsAg,DNA疫苗)、阻碍免疫的刹车(如PD-1/PD-L1Inhibitors)等,而靶向先天免疫的靶点目前是算主流应该属TLR(如TLR3和TLR7),本文会简单介绍一下比较新的STING和HBV之间的关系 。

Replication of HBV DNA, HBsAg production and HBeAg secretion[3]
STING (Stimulator ofinterferon genes)
正如其名,STING是一类干扰素IFN的刺激基因,也就是说能够上调IFN表达的那类基因,过去也被称为TMEM173,ERIS, MITA, MPYS 和 NET23。一般情况下真核细胞的DNA会固定的活动于细胞核内,只有发生意外状况时DNA才会涌入到细胞质中,比如细胞受损,开始凋亡,比如有外界病原(病毒、细菌等)的侵入,带入外源DNA片段等。这个时候细胞质中巡逻的DNA sensor就会及时发现,并启动相应的补救措施。这类DNAsensor一方面能够直接抵抗病毒或细菌的复制,一方面赶紧向大本营报信,直接激活机体的免疫能力,招来援军,很有全局意识。细胞会根据这些可靠的信号,通过不同通路的比较确认,最终决定选择哪种方式:比如通过NFκB激活下游的炎症因子,比如通过Caspase 1激活下游释放IL-1β等你细胞因子,比如经p62和NDP52的介导诱发自噬(autophagy),比如经Caspase 9诱发凋亡(apoptosis)或RIP3激活焦亡(necrosis)等同归于尽的方式,我想细胞在有余地的情况还是会选择另一条维护自我的通路——经STING-TBK1-IRF3诱导表达IFNs进行抵抗。[4]过去的研究曾多次表明细胞质中的DNA会被DNA sensor(DAI, IFIX, DDX41等)识别后将信号传递到 STING,再激活下游通路最终上调IFN的表达,增强先天免疫能力。直到2013年,一个课题组发现环状二核苷酸(cyclicdinucleotides, CDNs)可跳过DNA sensor(DDX41)直接激活STING通路;紧接着又有人发现cGAS可以将细胞质中的DNA转化成cGAMP来激活STING。[5, 6]沿着这两篇Science可以发现陈志坚教授(美国国家科学院院士)的课题组在这个领域是领军人物,他们连续发表许多关于STING通路的高水平文章。
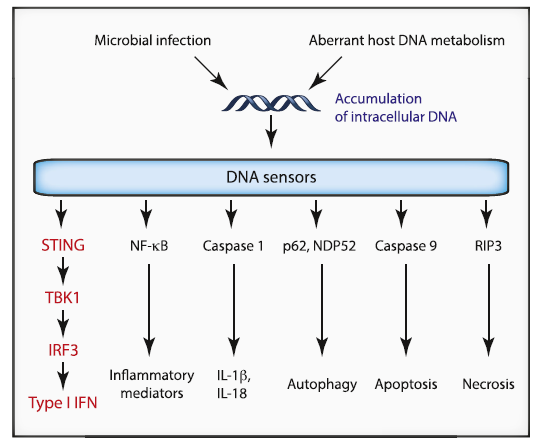
Cellular Functions Stimulated by DNA[4]
记得Invivogen的报告中提到过许多病毒的RNA或DNA可激活STING通路,比如(-)ssRNA的禽流感病毒(Avian influenza)、新城鸡瘟病毒(Newcastledisease virus)、仙台病毒(Sendai virus)和水疱性口炎印第安纳病毒(Vesicularstomatitis Indiana virus)等,再如(+)ssRNA的冠状病毒(Coronavirus)、登革热病毒(Dengue virus)、HCV, dsDNA类病毒也有报道,如腺病毒(Adenovirus)和人巨细胞病毒(Human cytomegalovirus)等,相比而言,对HIV的研究还是更多一些,比如人们发现HIV感染后会激活IRF3依赖的先天免疫,其中需要有活性的cGAS提供帮助,并且通过RNAi的筛选在human monocyte-derived DCs (MDDCs)中发现Polyglutaminebinding protein 1 (PQBP1)可以结合HIV反转录产生的cDNA,在与cGAS一同激活IRF3的通路。[7]也有人报道HIV可调用细胞的蛋白NLRX1(一种NLR蛋白)阻止STING与下游TBK1的相互作用。[8]
然而,STING对HBV的影响却鲜见报道。稍稍搜索一番,感觉在2015年之前通过调节先天免疫抑制HBV的文章还集中在TLR上,近两年也有人开始关注STING,我想用不了多久就会(可能已经)有人开展相关研究。
早期人们提出病原体相关分子模式(pathogen associated molecular patterns, PAMPs)以及模式识别受体(pattern recognition receptors, PRRs)的概念,最为常见的应该算TLR(Toll-like receptor)和RLR(RIG-I-like receptor),让人们更清晰的理解了细胞中调节先天免疫抵抗病原微生物在分子水平上的机理,这种逻辑也让让人们有机会发现更多的类似的机制。就好比通过病毒颗粒的结构,大致可以预测他们的感染方式、传播途径、存活条件以及如何抑制病毒的复制,也好比不论我们吃的是什么,被机体消化吸收的基本物质只有那些,最终要么变成氨基酸、要么核苷酸、要么葡萄糖等等。模式识别的方式,仅仅看一种病原体侵染细胞后会生成哪种固有的、保守的模序,接着会被细胞内哪些不同的受体识别、激活并将信号传递下去。
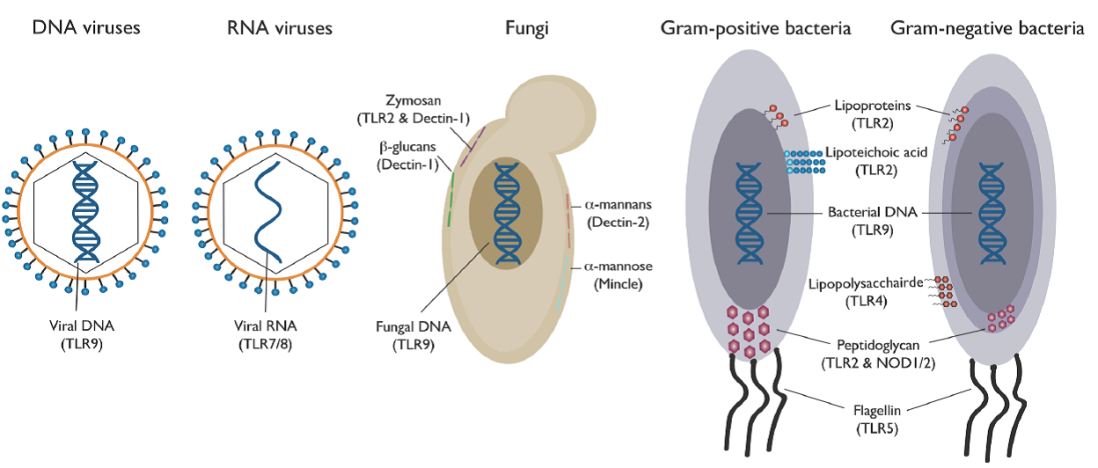
Recognition of PAMPs from different classes of microbial pathogens
对于HBV而言,肝脏中的巨噬细胞(macrophage)、树突细胞(dendritic cell)和肝非实质细胞(livernonparenchymal cells, NPCs)的TLRs会被相应配体激活,诱导表达I型IFNs(type I IFNs,如IFNα和IFNβ)、前炎性细胞活素(proinflammatorycytokine,如TNFα)和趋化因子(chemokine)等。I型IFNs会激活JAK-STAT通路,诱导IFN刺激基因(ISGs)的表达,可通过抑制HBV的cccDNA转录、基因组RNA包入衣壳而抑制HBV的复制;肝细胞中被活化的RLR同样可以诱导表达I型IFNs和前炎性细胞活素,也可以通过MAPK、NFκB、IRF3活化细胞内抗病毒通路,诱导表达抑制HBV复制的蛋白;T淋巴细胞和NK细胞被激活后可诱导表达II型IFNs(IFNγ),接着诱导IDO(indoleamine2, 3-dioxygenase)和APOBEC3G(apolipoproteinB mRNA-editing enzyme catalytic polypeptide 3 protein G)等抗病毒蛋白的表达,可抑制HBV的复制。[9]对于TLR而言,研究比较多的应属TLR3和TLR7了,目前已有多个案例进入临床试验。[10]

Molecular pathways of TLR- and RLR-mediated innate immunity against HBV [10]
如今,STING这颗耀眼的新星,站在TLR的肩膀上来到的人们眼前。既然有人报道她可以被cGAMP激活并诱导表达IFNs(与TLR7/8很像),自然就会有人重复与TLR类似的抗HBV现象,希望筛选到一种agonist能够激活这条通路,这也是一段反复验证的过程。
也是在2013年,就在cGAS-cGAMP-STING这条主线被揭示不久,人们发现血管阻断剂(Vascular disrupting agents)DMXAA(又称Vadimezan或ASA404,可破坏血管的生成,进而抑制肿瘤的生长,已进入PhaseII临床试验)可以直接与小鼠的STING结合后激活TBK1-IRF3信号通路,诱导IFNβ的表达,在人的PBMC和THP1细胞中却没有发现该现象。[11]大概在同一时间,另一个能早期发现能上调IFN表达的分子CMA(10-carboxymethyl-9-acridanone)也可激活STING通路,并且也是在小鼠的细胞中可见,人源细胞不可见。[12]一条新发现的通路,只要有合适的特异性的小分子抑制剂就比较容易着手研究这条通路的作用、成员、上下游信后传递关系等。终于,研究HBV的实验室等不及了,开始建立方法,筛选STING的agonists。
去年年初,一篇文章正式报道STING的agonists诱发先天性抗病毒免疫,可抑制HBV的复制。文中利用鼠巨噬细胞和肝细胞共培养的系统模拟肝中的微环境,利用参照药物DMXAA刺激,结果发现巨噬细胞可释放大量的细胞因子,可明显减少鼠肝细胞中HBV的核衣壳蛋白量。同时他们还比较了TLR通路与STING通路激活后细胞做出的不同反应(用TLR特异的agonists容易实现,如TLR1/2的Pam3CSK4,TLR3的poly(I:C), TLR4 的LPS和TLR7的Gardiquimod等等),结果发现TLR活化后的细胞多数分泌大量的炎症因子或趋化因子,而STING活化后主要产生IFN,进而促使更多的ISGs表达,抑制HBV的复制。[13]
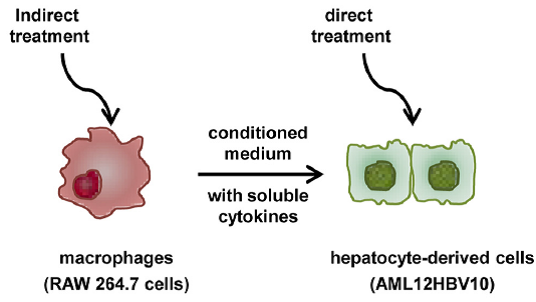
Suppression of HBV replication in hepatocytes by TLRresponse activation in macrophages[13]
一般情况下,HBV没有合适的动物模型,目前已知HBV可感染人和一些灵长类动物比如黑猩猩,可对小鼠和大鼠这类实验室常用的动物却没有感染力,目前常用的动物模型除灵长类动物之外,有被WHV(woodchuck hepatitis virus)感染的Woodchuck(土拨鼠),也有被DHBV(duck hepatitis B virus)感染的Duck。用于药物开发用的动物模型受限,也是限制HBV药物研制的原因之一。现在看来,HBV的感染或复制的能力不强,或许是容易形成慢性肝炎的因素之一。有数据显示95%的成人感染HBV后会再6个月内通过免疫能力清除体内的HBV,90%的婴儿和30%的1~5岁的儿童感染HBV后易形成慢性肝炎。
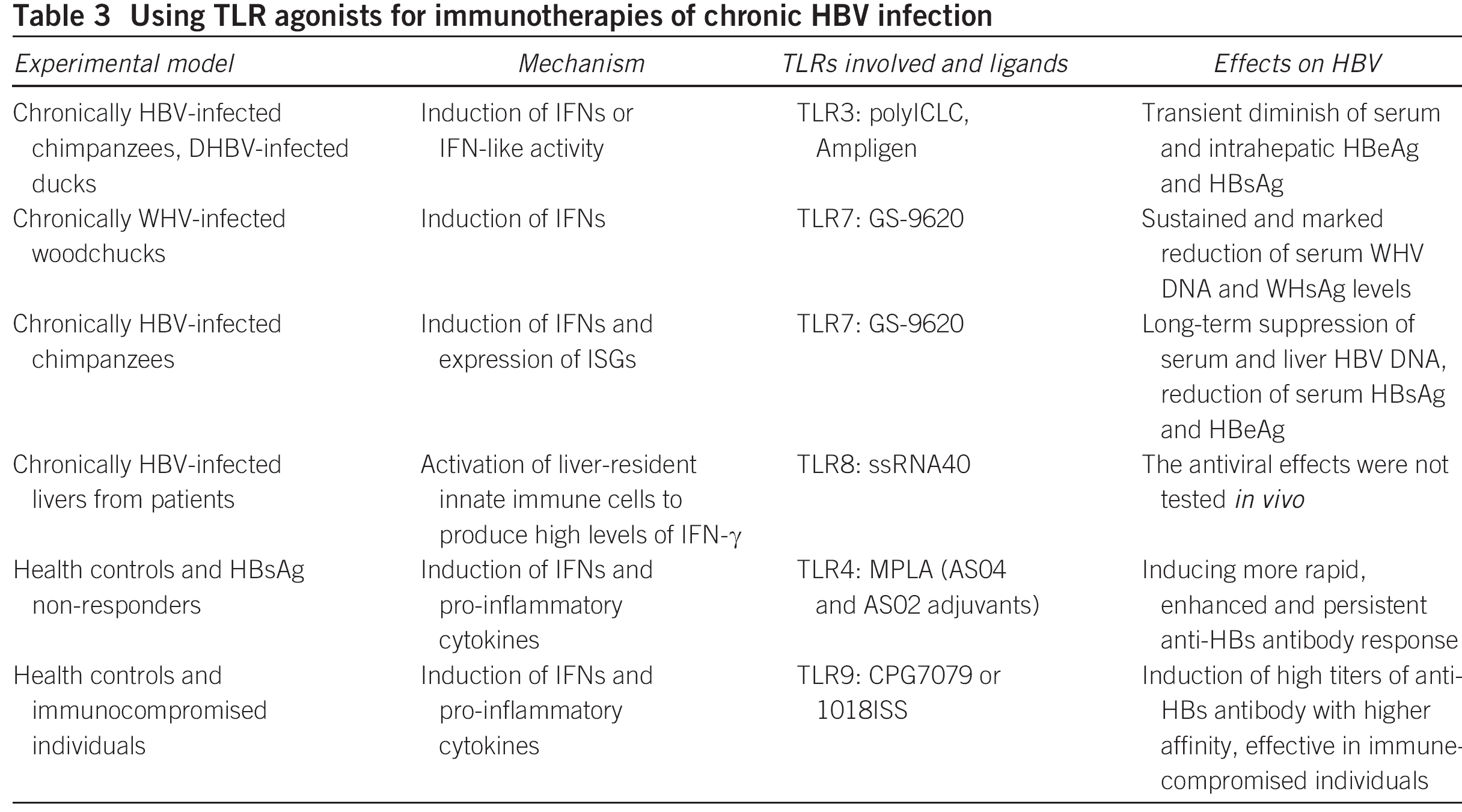
DHBV, duck hepatitis B virus; MPLA, monophosphoryl lipid A; polyICLC,polyriboinosinic : polyribocytidylic acid; PWH, primary woodchuck hepatocyte [10]
与此同时,有文章报道了HBV拮抗STING的一种方式:破坏STING在K63位的泛素化修饰。(这类研究在HIV的基础研究中常见,一旦发现一个新的抗病毒蛋白,不久之后就会有报道显示病毒有特有的方式可以拮抗,否则这类病毒将会毁灭。)文中利用瞬转HBV感染性克隆的Huh7细胞、稳定产生HBV的HepAD38细胞、HBV感染的HepaRG细胞核原代人肝细胞等细胞株观察到STING通路诱导表达的IFNβ可明显的抑制病毒的复制;同时作者在过表达STING的细胞株中也发现随着HBV感染剂量的增加,IFNβ的分泌成剂量依赖性减少,通过逐步排查,发现时病毒的聚合酶(polymerase)干扰了STING的信号通路。再缩小范围,分析比较了其中的几个亚基或结构域:末端蛋白(terminal protein, TP), 间隔序列(spacer),反转录酶(reversetranscriptase, RT)和 RNase H (RH),发现RT/RT结构域可与STING直接结合,后来作者比较分析了TING活性的泛素化修饰,才揭开K63的重要性。[14]
后来陆续也有几篇文章报道了HBV和STING之间的博弈关系,在不同的细胞中、不同的实验方法和技术、不同的ISGs、不同的in vitro和in vivo的模型,反复的验证HBV与STING的交叉点并提出cGAS-STING---IFNs这条通路可能成为新的医药研发的靶点,治疗乙肝的新策略,或许真的会用在日后的联合治疗中。

Innate immune signaling pathway of varies nucleic acidsensor interfered by the HBV viral proteins[15]
回过头来再看,STING只是众多DNA sensor通路中的一条,其实细胞有很多种武器或部队可以用来识别、攻击HBV,只是有时敌人过于顽强、善于隐秘,难以被清除而已。在HBV感染期间,会产生多种核酸类中间产物,如单链RNA(ssRNA)、双链的RC-DNA、cccDNA、双链的RNA(dsRNA)等。细胞质的DNA受体(如cGAS、DDX41)识别DNA后激活ER结合的STING-TBK1-IRF3通路,诱导IFNs表达;RLR(如RIG-I,MDA5)结合细胞之内RNA后会通过TBK1-IRF3激活结合线粒体的MAVS下游通路;4种内体中的TLR(3,7/8和9)可分别识别dsRNA、ssRNA和无乙酰化的CpGDNA,激活TICAM-1和MyD88依赖的通路上调I型IFN。[15]
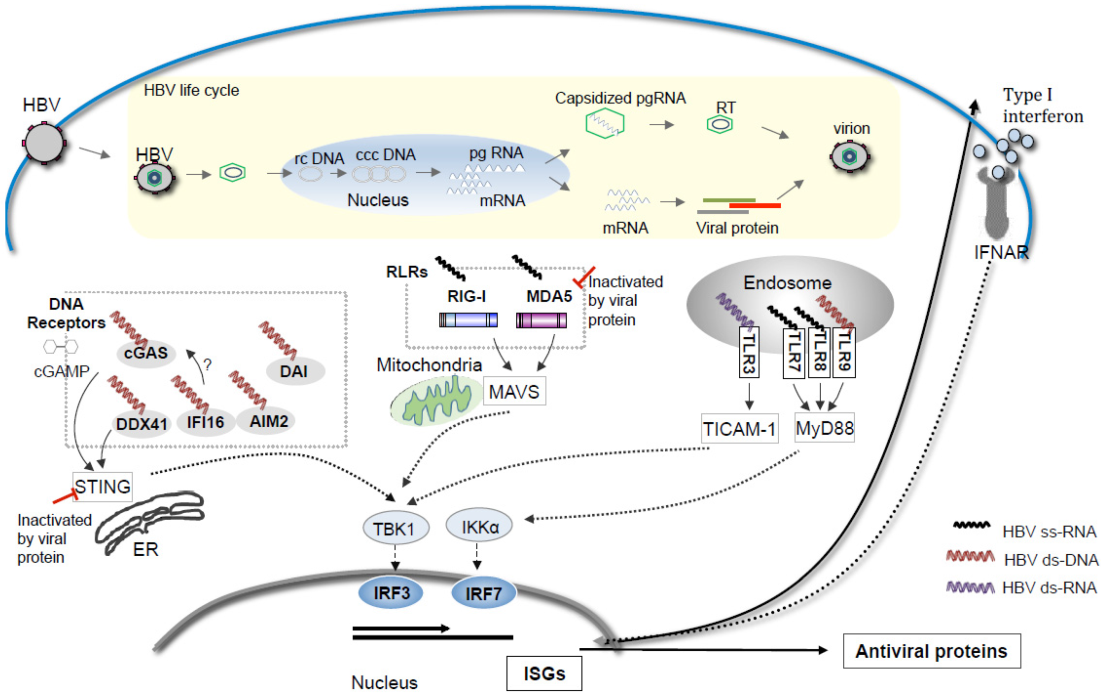
Possible Nucleic acid sensors participation in HBV pattern sensing in thelife cycle of HBV[15]
希望中国的科学研究能在“十三五”期间登上新的台阶,希望国内的科研单位、制药单位能在一些创新型领域占一席之地,希望人们早日获得治愈乙肝的良药!
TJ泰河园
Aug 28, 2016 Sunday
Reference
4. Paludan, S.R. and A.G. Bowie, Immune sensing of DNA. Immunity, 2013. 38(5): p. 870-80.
https://wap.sciencenet.cn/blog-571539-1001250.html
上一篇:关于Video的那些事
下一篇:Discovery of cGAS-STING pathway