博文
脑内伽马事件的意义【自然】  精选
精选
|
美国耶鲁大学科学家发现,只要向小鼠大脑“播放”一段特定的“伽马”节律,就能成功“欺骗”它们,让它们以为自己看到了本不存在的视觉信号!这一惊人发现,彻底改写了我们对大脑如何处理信息的理解。这项研究的真正意义在于,它为我们“破译”感官知觉乃至意识的编码方式提供了全新的视角,并可能为阿尔茨海默病、精神分裂症等众多与脑节律紊乱相关的重大疾病,带来可解读的早期生物标志物。这篇重磅论文2025年10月8日刚刚发表在国际顶尖学术期刊《自然》(Nature)上。
Flexible perceptual encoding by discrete gamma events | Nature
行为背后的认知过程与新皮层中特定的时空神经活动模式相关[1-6]。这些模式源于同步突触活动[7],常被作为振荡活动进行分析,但也可能表现出难以被有效检测的非周期性动态。本研究开发了一种新颖的分析方法,可将模式化活动分解为离散的网络事件,并运用该方法追踪小鼠视觉皮层(V1区)的伽马活动(30-80Hz)。研究发现,伽马事件发生率随觉醒状态变化,单个事件既可能在短暂的振荡发作中成簇出现,也可能单独发生。单个事件能同步跨皮层层的神经放电,并促进视觉编码的增强。V1区的伽马事件由背外侧膝状体核(dLGN)的模式化输入诱发,且会被dLGN的光遗传学调控所抑制,这表明伽马事件有助于视觉信息的丘脑-皮层整合。在行为学实验中,小鼠出现视觉提示的行为反应前,伽马事件发生率会稳步上升,且能预测逐次实验的表现。抑制V1区伽马事件会损害视觉检测能力,而诱发伽马事件则会引发行为反应。伽马事件与行为之间的这种关联具有感觉模态特异性,并会随任务目标的变化而快速调节。因此,伽马事件可根据行为情境,为视觉信息提供灵活的编码方式。
正文
新皮层的神经活动呈现出复杂的时空模式,这些模式能动态反映行为状态和任务参与度的变化[4,8-12]。此外,活动模式异常是多种神经发育障碍和精神疾病的典型特征[13,14]。皮层局部场电位(LFP)中的活动模式主要源于局部神经回路中的突触电流[7]。高频活动(尤其是30-80Hz的伽马波段)与注意力、感知觉和记忆等认知过程相关[15,16]。通常认为,这类活动对应于振荡过程[4,8,9,17,18],该过程由兴奋性神经元与抑制性神经元之间的局部相互作用产生[8,9,19-23],并可“调控”传入信号[8,24,25],从而促进信号向输出结构传递[24,26]。然而,这一理论框架无法解释关于皮层活动的部分关键观测结果[25,27-31],且局部振荡器支持脑区间信息交流的机制仍不明确[32-34]。
相反,模式化皮层活动可能代表着重复出现的单个网络事件,而这些事件难以通过标准方法精确且可靠地检测。此外,模式化活动的发作也可能反映皮层各层之间的动态相互作用[35],但其空间结构却很少被量化。因此,要建立皮层活动模式与行为之间的全面关联,需要开发新的方法,以在动态调节的皮层活动中,检测并量化具有一致时空特征的离散网络事件。
为验证模式化皮层活动是否代表一系列离散的网络事件,我们在头部固定、可自由跑动的小鼠V1区跨皮层层记录了局部场电位(图1a、b),并开发了一种基于状态和频谱-时间特征对限带活动进行聚类的分析方法(CBASS;方法部分、扩展数据图1及补充信息)。CBASS整合了以往研究的思路,旨在识别跨通道具有一致动态特征的重复活动模式[36,37],同时专门分析特定频段内的单个周期[31,38]。关键在于,CBASS以单个事件为分析单位,因此能够检测出周期性各异的神经过程[29]——从分散的单个事件到短暂或持续的振荡[28,39,40]。该方法首先分离出参考通道中特定频段内具有能量的候选事件,随后保留那些跨通道层间和时间特征一致、且在特定目标状态下富集的事件(方法部分、图1d及扩展数据图1)。
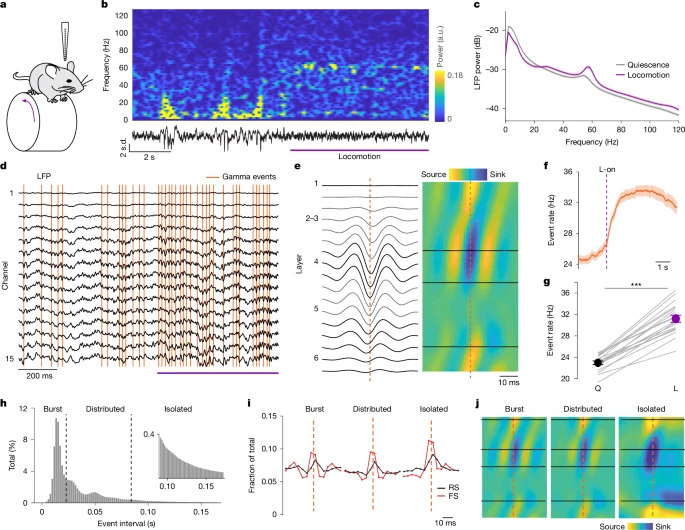
图1:CBASS方法将伽马波段(30-80Hz)活动的状态依赖性变化与特定网络事件关联
a. 头部固定小鼠在跑轮上进行V1区跨皮层层记录的示意图。
b. 示例数据:显示一个局部场电位通道的信号及其短时傅里叶变换(从静息状态向运动状态过渡,紫色标注)。a.u.,任意单位。
c. 跨通道局部场电位平均功率(n=19只小鼠),显示运动状态下伽马波段(30-80Hz)功率选择性升高。
d. 对运动状态下V1区数据应用CBASS方法:多通道局部场电位(黑色)经伽马波段(30-80Hz)滤波;在参考通道的滤波信号谷值处筛选候选事件,保留那些跨通道相位和振幅特征可预测运动状态的事件(橙色柱)。
e. 伽马事件前后的平均局部场电位(左)及相关的电流源密度(CSD)图谱(右)。事件与活动从第4层向表层皮层传播、随后向深层皮层传播相关。橙色虚线标记伽马事件。
f. 运动开始(L-on)前后CBASS检测到的伽马事件发生率(n=19只小鼠)。
g. 运动状态(L)下事件发生率升高(n=19只小鼠)。Q,静息状态。
h. 伽马事件的事件间间隔平均分布。事件分为三组:(1)成簇事件,相邻事件间隔≤27.3ms(1.5个平均周期);(2)分散事件,相邻事件间隔>27.3ms且≤90.9ms(5个平均周期);(3)单独事件,相邻事件间隔>90.9ms(n=19只小鼠)。图中还展示了“单独事件”部分的放大视图(插图)。黑色虚线表示组间分界。
i. 跨皮层层胞外记录的规则放电(RS;黑色;n=457个单位)和快速放电(FS;红色;n=260个单位)神经元在成簇、分散和单独伽马事件前后的群体平均放电分布。橙色虚线标记伽马事件。
j. 成簇、分散和单独伽马事件的平均电流源密度图谱(n=19只小鼠)。橙色虚线标记伽马事件。
误差棒表示标准误(s.e.m);阴影区域表示平均值±标准误。统计学显著性:*P≤0.05,***P≤0.001。详细统计数据参见补充表1,统计样本参见补充表2。
已有研究表明,小鼠运动时V1区伽马波段(30-80Hz)功率会选择性升高[4,11,17](图1b、c),这为研究行为状态下动物离散、重复的皮层网络事件提供了明确场景。CBASS方法在清醒小鼠V1区检测到持续发生的伽马事件(平均发生率为24.3Hz;n=19只小鼠)。在频谱和通道协方差与本研究记录数据匹配的随机化数据中,事件检测的可能性极低(扩展数据图1i-k),但更换参考通道后检测结果仍稳定可靠(扩展数据图1n-o)。通过基于振幅阈值的更简单方法也能检测到伽马事件(扩展数据图1p-q),这表明此类事件是清醒状态下皮层活动的固有组成部分。然而,CBASS方法通过要求跨皮层层的一致动态特征,实现了更灵敏的检测。综上,这些结果表明CBASS方法可稳定检测单周期网络事件。所有统计检验和样本分别详见补充表1和补充表2。
伽马事件的发生频率随时间存在显著变化(图1d)。尽管如此,在不同行为状态、视觉刺激条件及瞬时发生率变化下,伽马事件的平均局部场电位(LFP)和电流源密度(CSD)图谱仍保持稳定(图1e、h-j及扩展数据图2v-ab)。伽马事件周围的平均场电位在伽马波段具有能量(扩展数据图2v),且在伽马事件高发期,伽马波段的LFP功率会升高(扩展数据图2g)。运动状态下伽马事件发生率升高(图1f、g及扩展数据图2k、l),且与作为觉醒状态生物标志物的瞳孔直径相关(扩展数据图2m)。伽马事件既会以振荡成簇的形式出现,也会以单独事件的形式存在(图1d、h)。无论是单独发生、分散成组发生还是紧密成簇发生的伽马事件,在牵引V1区神经元放电方面的效能均相似(图1i),且具有一致的CSD图谱(图1j)。不过,伽马事件对LFP频谱的影响取决于其瞬时发生率(扩展数据图2z-ab):单独事件会诱发宽频带(30-80Hz)的能量,而成组事件则诱发较窄频带(约55Hz)的能量[17]。这表明,LFP中窄带和宽带伽马功率的活动时期[17,29,41],实则是同一网络机制产生的不同结果。
除了与运动相关的伽马活动外,小鼠V1区还存在其他显著的模式化活动模式,包括受视觉调控的强效β波或低伽马振荡(15-30Hz,以下简称β波)[6,42,43]。通过CBASS方法检测到的β波事件受视觉刺激调控(扩展数据图2c、d、f、n-q),且与伽马事件具有不同的层间分布图谱——β波事件对深层皮层的激活更强(扩展数据图2r、s)。运动状态或瞳孔直径对β波事件发生率的调控作用不显著(扩展数据图2k-m)。β波事件与伽马事件在快速时间尺度上交替出现,表明网络过程可快速切换(扩展数据图2a、b)。此外,伽马事件与β波事件的共标记概率高于随机水平(扩展数据图2j),且共标记事件具有介于两者之间的图谱特征(扩展数据图2s-u)。与近期提出的模型[6,42,43,44]一致,伽马事件与β波事件可能分别涉及不同但相互作用的兴奋性-抑制性局部环路机制。
以往研究认为,皮层伽马活动要么是由兴奋性与抑制性神经元间的相互作用在局部产生[8,9,19,20],要么是从丘脑等传入结构中被动继承而来[17,45]。与此不同,本研究发现V1区单个伽马事件与活动从第4层向第2-3层和第5层的强效传播相关(图1e、j及扩展数据图2v-ab),这与皮层环路对丘脑驱动信号进行主动前馈整合的已知特征相符[46,47]。为进一步验证皮层伽马事件是否由丘脑-皮层输入产生,我们在背外侧膝状体核(dLGN)的背部区域表达了通道视紫红质-2(ChR2)[8,48](方法部分、图2a及扩展数据图3a、b)。通过模式化的1毫秒光脉冲序列激活V1区中的dLGN末梢,结果显示:dLGN末梢刺激诱发的皮层反应其CSD图谱与伽马事件的CSD图谱具有显著的余弦相似度(扩展数据图3s、t),且在不同刺激模式下均保持一致(扩展数据图3u-w)。规律的dLGN刺激序列会产生非生理性的梳状频谱,而泊松分布的刺激序列则会导致全频段功率升高[29](图2d及扩展数据图3p、q)。相比之下,还原V1区记录到的伽马事件自然时间分布的脉冲序列(图1d、h及扩展数据图2i),无论在何种行为状态下,均能选择性诱发皮层伽马活动的宽带(30-80Hz)和窄带(约55Hz)成分升高(图2d、e及扩展数据图3o-r、x-ac)。由此可见,V1区伽马活动的频率特征在很大程度上可由伽马事件的时间分布解释,且可通过简单模型复现[29](扩展数据图3g-n)。随后,我们在丘脑网状核(TRN)中表达生长抑素(SST)的抑制性神经元中表达ChR2,并在dLGN上方植入光纤(见方法部分;图2f及扩展数据图3c-f)。通过激活SST+ TRN末梢来调控dLGN活动[49,50],结果显示:在静息和运动状态下,V1区伽马事件的发生率均显著降低(图2h-j),且整体伽马功率也中度下降(图2g)(扩展数据图3ad-aj)。因此,V1区伽马活动既非仅由局部环路产生,也非从dLGN以振荡形式继承而来,而是通过皮层各层对dLGN输入信号的主动整合形成[51,52]。
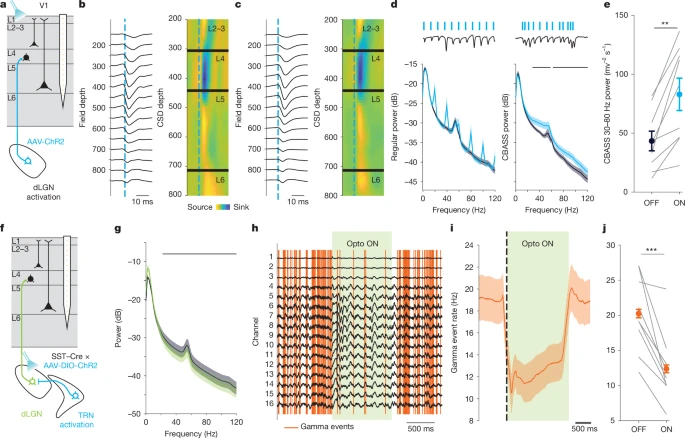
图2:皮层伽马事件由丘脑输入驱动
a. 头部固定于跑轮上的小鼠在进行V1区跨皮层层记录的同时,对dLGN末梢进行光遗传学激活的示意图。实验前5-8周,向小鼠dLGN内注射AAV5-hSyn-hChR2-eYFP病毒。
b. 示例小鼠中,低强度激活V1区dLGN末梢(474nm波长,1-5mW/mm²功率,持续1毫秒)所诱发的平均LFP及相关CSD图谱。蓝色虚线标记光脉冲时刻。
c. 多只小鼠(n=9只)的结果与b一致。dLGN末梢激活诱发的反应,其层间分布图谱和时间进程与伽马事件相符。
d. 跨通道平均LFP功率:dLGN激活前(深蓝色)与激活期间(浅蓝色)。规律脉冲序列(左图,20Hz,n=4只小鼠)诱发梳状频谱;而还原伽马事件自然时间分布的刺激序列(右图,n=9只小鼠)则诱发V1区伽马活动的宽带(30-80Hz)和窄带(约55Hz)成分升高。
e. CBASS方法定义的dLGN分散刺激序列作用前(深蓝色)与作用期间(浅蓝色),伽马波段(30-80Hz)的平均功率(n=9只小鼠)。
f. 头部固定于跑轮上的小鼠在进行V1区跨皮层层记录的同时,通过光遗传学激活SST+ TRN神经元的抑制性末梢以调控dLGN活动(474nm波长,连续光刺激,50mW/mm²功率)的示意图。实验前5-8周,向SST-Cre+小鼠的TRN内注射AAV5-DIO-hChR2-eYFP病毒,并在dLGN上方植入光纤。
g. 光刺激前(灰色)与刺激期间(绿色)的跨通道平均LFP功率(n=9只小鼠)。
h. 示例LFP记录:静息状态下,dLGN光刺激(绿色标注)期间V1区伽马事件(橙色柱)发生率降低。Opto,光遗传学。
i. dLGN光刺激前后,V1区CBASS方法检测到的伽马事件发生率(n=9只小鼠)。黑色虚线标记光脉冲开始时刻。
j. dLGN光刺激期间,V1区伽马波段(30-80Hz)的平均功率(n=9只小鼠)。
误差棒表示标准误(s.e.m);阴影区域表示平均值±标准误;黑色水平条表示统计学显著性:*P≤0.05,***P≤0.001。详细统计数据参见补充表1,统计样本参见补充表2。
为探究网络伽马事件对单个神经元的影响,我们在清醒小鼠的静息和运动状态下,对皮层第2-5层进行全细胞膜片钳记录,同时监测跨皮层层的LFP(图3a、b及扩展数据图4)。结果显示:伽马事件的发生与皮层神经元的去极化“上状态”(up-state)相关(扩展数据图4t),且与膜电位(Vm)的快速偏转同步[21,41](图3b、c及扩展数据图4c、d、i、j、o、p)。伽马事件发生时,全频段的Vm功率均升高(扩展数据图4e、k、q),且所有皮层层中Vm与LFP在伽马波段的相干性均选择性增强(图3d、e及扩展数据图4f、l、r)。在所有皮层层中,伽马事件的发生与神经元放电时刻精准同步;且在细胞内记录(扩展数据图4g、m、s、u、v)和细胞外记录(图3f、g及扩展数据图5e-n)中,均观察到伽马事件与“放电-LFP同步性”显著增强相关。伽马事件相关的放电在第4层发生最早,在第2-3层发生最晚,这与丘脑-皮层前馈加工的特征一致(图3f)。第2-3层的同步性最强(图3g及扩展数据图4v、5k-n);且与推测为兴奋性神经元的规则放电(RS)单元相比,推测为抑制性神经元的快速放电(FS)单元其同步性显著更高(扩展数据图5l、o),这与以往研究报道[8,9,53]高度一致。伽马事件对规则放电单元和快速放电单元均具有强效牵引作用,这进一步表明:这些网络事件由dLGN驱动,但会激活皮层固有的共振局部环路[8,9]——该环路可能对伽马波段的信号进行选择性锐化和传播[46,47]。
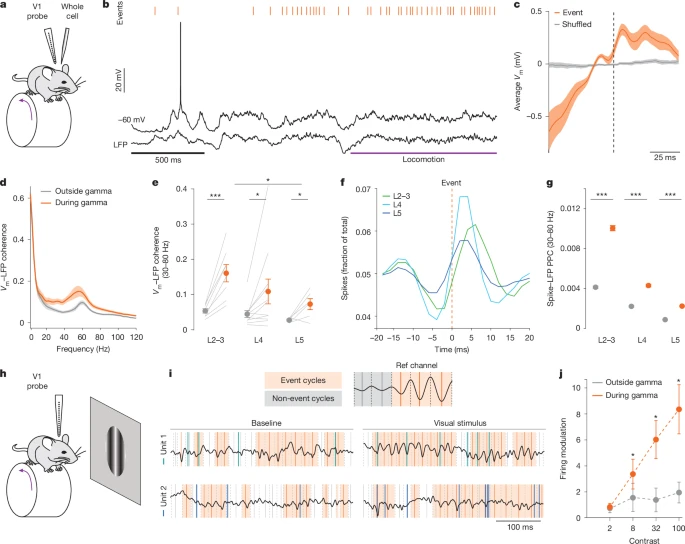
图3:伽马事件调控神经元放电时刻并增强视觉编码
a. V1区全细胞膜片钳记录与跨皮层层记录同步进行的示意图。
b. 第4层神经元的膜电位、反转的LFP信号,以及运动开始(紫色标注)前后的伽马事件(橙色标注)。
c. 伽马事件前后的平均膜电位(n=25个神经元)。黑色虚线标记伽马事件发生时刻。
d. 伽马事件周期内(橙色)与周期外(灰色)Vm与LFP的相干频谱(n=25个神经元)。
e. 伽马事件周期内(橙色)与周期外(灰色)的整体伽马相干性(30-80Hz)(第2-3层8个神经元,第4层11个神经元,第5层6个神经元)。
f. 细胞外记录到的规则放电单元在伽马事件前后的群体平均放电分布:第2-3层(绿色)、第4层(青色)、第5层(深蓝色)。
g. 规则放电单元在伽马波段(30-80Hz)的整体放电-LFP成对相位一致性(PPC):伽马事件周期内(橙色)与周期外(灰色)(第2-3层82个单元,第4层69个单元,第5层281个单元)。
h. 视网膜拓扑定位视觉刺激呈现期间进行跨皮层层记录的示意图。
i. 伽马事件周期内与周期外神经元放电分析示意图(上图);两个V1区规则放电单元在刺激开始前后的活动示例,展示伽马事件周期内的视觉诱发放电(下图)。
j. 伽马事件周期内(橙色)与周期外(灰色),规则放电单元对不同对比度光栅刺激的放电反应调制(n=47个规则放电单元)。
误差棒表示标准误(s.e.m);阴影区域表示平均值±标准误。统计学显著性:*P≤0.05,***P≤0.001。详细统计数据参见补充表1,统计样本参见补充表2。
尽管伽马活动与认知过程的整体关联已得到充分证实,但伽马事件在皮层感觉加工中的具体作用仍不明确。视觉刺激会显著增强伽马事件周期内的放电-LFP同步性(扩展数据图5f、l、o)。在自发活动期间,伽马事件的发生与规则放电单元的放电无相关性;但在高对比度漂移光栅刺激呈现期间,二者则出现相关性(扩展数据图6a-f),这表明视觉诱发的放电更倾向于在伽马事件期间发生。因此,我们对伽马事件周期内与周期外的视觉反应进行了分析(图3h、i)。结果发现:在伽马事件周期外,视觉刺激对规则放电单元的放电几乎无调制作用(图3j及扩展数据图6i);而在伽马事件周期内,无论处于何种行为状态,视觉诱发的放电均显著增强(图3j及扩展数据图6i、k、m)。部分视觉诱发活动也会在β波事件期间发生,但选择性较低(扩展数据图6g、h)。由此可见,伽马事件可对皮层神经元的视觉诱发放电进行整合。
为验证伽马事件对视觉编码的增强作用是否有助于感觉引导行为,我们训练小鼠完成视觉对比度检测任务(方法部分及图4a)——该任务的表现具有行为状态依赖性(扩展数据图7a-g),且依赖V1区的功能(扩展数据图7h-l)。采用可降低皮层伽马事件的刺激方案(图2f-j)调控dLGN活动,会导致小鼠的视觉检测表现显著下降(图4b-e)。相反,对训练后的小鼠进行dLGN末梢的模式化激活,会诱发其行为反应(表现为假阳性试次增多)(扩展数据图7m-r)。该效应对刺激强度敏感,且在还原CBASS方法检测到的事件自然模式时,效应最为显著(扩展数据图7q-r)。综上,这些数据表明:V1区伽马事件有助于视觉信息整合,且与下游行为反应的启动相关。
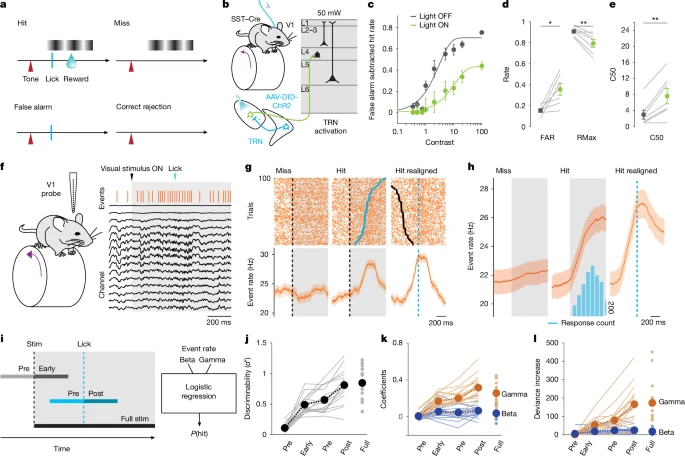
图4:伽马事件可预测视觉检测任务中的行为反应,抑制伽马事件会损害任务表现
a. 视觉检测任务的试次类型示意图。试次开始以声音信号提示:若呈现光栅刺激,小鼠舔舐可获得水奖励(击中试次,hit);无视觉刺激时舔舐(假阳性试次,false alarm)会导致惩罚期(time-out);刺激呈现时未反应(漏报试次,miss)或无刺激时未反应(正确拒斥试次,correct rejection)均无结果。
b. 向丘脑网状核(TRN)注射AAV5-DIO-hChR2-eYFP病毒的头部固定SST-Cre+小鼠(n=7只)执行a中的任务。在随机穿插的试次(30%)中,通过光纤向背外侧膝状体核(dLGN)上方施加蓝光(570nm波长,50mW/mm²功率,连续刺激),以激活SST+ TRN抑制性末梢。
c. 常规试次(灰色)与TRN激活试次(绿色)中,“假阳性率校正后的击中率”(平均值±标准误)随刺激对比度的变化。对两种条件下的击中率分别拟合S型函数(n=7只小鼠)。结果显示TRN激活会降低检测表现。
d. 最大对比度刺激下,常规试次(灰色)与视觉皮层(V1)失活试次(绿色)的假阳性率(FAR)与击中率(RMax)。TRN激活会升高假阳性率并降低最大击中率(n=7只小鼠)。
e. 常规试次(灰色)与V1失活试次(绿色)中,击中率达到50%时的刺激对比度(C50)。TRN激活会升高C50(n=7只小鼠)。
f. 任务执行期间的慢性记录示例:1个击中试次中的局部场电位(LFP)、伽马事件(橙色柱)、视觉刺激(灰色标注)及正确舔舐反应(蓝色箭头)。
g. 随机选取100个试次的伽马事件发生 raster 图(上图)及试次间平均事件发生率(下图):左侧为漏报试次,中间为击中试次,均以刺激开始时刻(对比度<7.5%;黑色虚线及标记)对齐;右侧为击中试次,以舔舐反应时刻(蓝色虚线及标记)对齐。
h. 任务试次中的群体平均伽马事件发生率(n=16只小鼠)。
i. 用于预测试次结果的逻辑回归分析窗口示意图:刺激早期(stim onset后300ms)、全刺激期(full visual stimulation)、舔舐前(反应前300ms或拒斥试次的平均反应前300ms)、刺激前(stim onset前300ms)、舔舐后(反应后300ms或拒斥试次的平均反应后300ms)。
j. 回归模型的敏感性(d′)在反应时刻前升高,且在反应后立即达到最高(n=16只小鼠)。
k. 伽马事件(橙色)与β波事件(蓝色)的模型系数(n=16只小鼠)。
l. 移除伽马事件与β波事件参数后的偏差增量(n=16只小鼠)。
为进一步明确伽马事件与视觉检测行为的关联,我们通过慢性植入的层间电极阵列,记录了小鼠执行任务期间的V1区活动(图4f)。结果显示:在击中试次(而非漏报试次)中,伽马事件发生率从刺激开始后持续上升,并在舔舐反应开始时达到峰值(图4g、h及扩展数据图8b、g);而β波事件发生率不受试次结果影响(扩展数据图8d-f、h)。整个试次期间,伽马事件的层间分布图谱及其对皮层神经元的牵引作用均保持稳定(扩展数据图9b-m)。我们采用逻辑回归模型,通过刺激与反应开始前后特定时间窗口内的伽马/β波事件发生率预测行为反应(图4i)。结果显示,随着小鼠接近舔舐反应时刻,预测准确率逐渐升高;偏差增量、参数随机化及系数值均表明,伽马事件发生率是预测逐次试次行为结果的关键因素(图4j-l及扩展数据图9n-w)。仅对静息状态下的试次进行分析时,上述结果仍成立,表明该关联并非由运动相关伽马事件驱动(扩展数据图9x-ab)。此外,胡须抖动(whisking)不会调控伽马事件发生率,进一步说明伽马事件与行为的关联并非简单由运动行为导致(扩展数据图8j-m)。假阳性试次中,伽马事件发生率升高及模型预测的显著性同样存在(扩展数据图8c、g及9s-w)。因此,伽马事件发生率升高可预示与任务相关的行为反应。
为验证行为反应前伽马事件发生率升高是否与“预期获得奖励”相关,我们训练无任务经验的小鼠在观看灰色屏幕时自由获取奖励(图5a)。结果显示:无论是否获得奖励,舔舐反应前的伽马事件发生率均无显著升高(图5b、d-g),表明伽马事件并不编码通用运动反应或奖励信号。为探究伽马事件是否可能代表“视觉刺激与奖励的习得关联”,我们将小鼠转入新实验范式:仅当舔舐反应在视觉刺激期间发生时,才可获得奖励(图5a)。该范式下,伽马事件发生率在行为反应前选择性升高(图5b、d-g及扩展数据图10c、d);这种调控作用起效迅速(视觉范式第一天即出现),且与运动或觉醒状态无关(图5c、e及扩展数据图10b-f)。随后将小鼠转回“自由奖励范式”,伽马事件与行为的关联立即消失(图5b、d-g)。
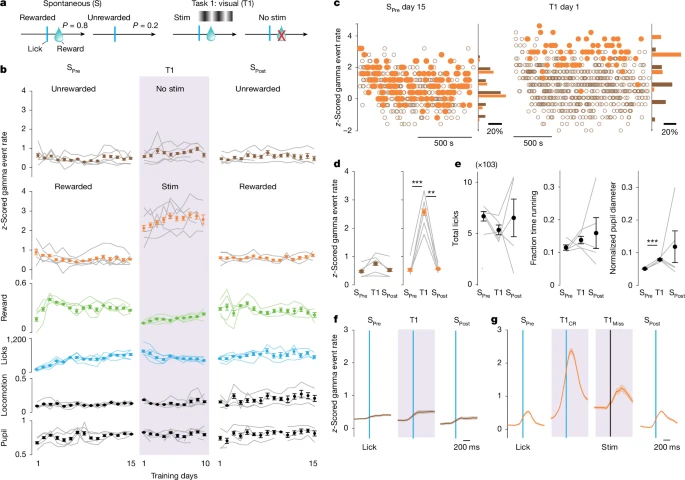
图5:任务情境变化对伽马事件发生率的快速调控
a. “自由奖励范式”(S)与“任务1(T1)”的试次类型示意图:首先训练小鼠15天,使其可自由获取奖励(SPre阶段);随后转入T1阶段(10天),仅视觉刺激期间舔舐可获奖励;最后转回自由奖励范式(SPost阶段,15天)。全程通过慢性植入电极记录V1区跨皮层层活动。
b. 从上至下依次为:无奖励舔舐(棕色)与有奖励舔舐(橙色)后300ms内的标准化伽马事件发生率、有奖励试次占比(绿色)、舔舐次数(蓝色)、运动时间占比(黑色)、瞳孔直径(黑色),数据按训练天数展示(n=5只小鼠)。
c. 示例小鼠在SPre阶段第15天与T1阶段第1天,无奖励试次(棕色)与有奖励试次(橙色)的伽马事件发生率。
d. SPre、T1、SPost范式中,无奖励反应(左)与有奖励反应(右)后300ms内的标准化伽马事件发生率(n=5只小鼠)。
e. SPre、T1、SPost范式中的总舔舐次数(左)、运动时间占比(中)、平均标准化瞳孔直径(右)(n=5只小鼠)。
f. SPre、T1、SPost范式中,无奖励舔舐前后的标准化伽马事件发生率(n=5只小鼠,蓝色)。
g. SPre、T1、SPost范式中,有奖励舔舐(蓝色)或刺激开始(黑色)前后的标准化伽马事件发生率(n=5只小鼠)。视觉提示诱发的反应对伽马事件发生率的提升作用,强于单纯视觉刺激。
误差棒表示标准误(s.e.m);阴影区域表示平均值±标准误。统计学显著性:*P≤0.05,**P≤0.01,***P≤0.001。详细统计数据参见补充表1,统计样本参见补充表2。
视觉关联条件变化对伽马事件发生率的“快速调控”,与强化学习的特征不符——后者的关联建立与消失需随时间逐步发生[54,55]。由于伽马事件与视觉整合相关,这种快速调控更可能表明:整体情境会驱动特定变化,使V1区在“视觉引导行为期间”的信息加工得到增强。这一推测得到以下结果支持:与“主动反应获得奖励”相比,“视觉刺激自动伴随奖励”仅能轻微提升伽马事件发生率(扩展数据图10a、g、h);此外,当小鼠接受听觉任务训练时,正确反应前V1区伽马事件发生率无升高(扩展数据图10a、i、j),表明非特异性整体觉醒或任务参与度并非伽马事件发生率快速调控的原因。因此,行为反应时的伽马事件发生率升高具有“感觉模态特异性”,且对任务情境敏感——仅当视觉信息被用于引导行为输出时,V1区才会出现这一现象。
综上,本研究结果为“皮层活动模式分析”提供了一种新思路,突破了以往以“振荡”为核心的主流分析框架局限。借助多通道记录技术,我们发现:将皮层活动模式视为“具有特征性层间结构的离散事件序列”,能有效捕捉小鼠V1区伽马活动的精细时间动态。尽管伽马事件常以紧密振荡成簇的形式出现,但也会以更不规则的间隔单独发生;这种复杂的时间分布特征,正是其独特功率谱特征的来源。无论是单独还是成簇的伽马事件,均能调控单个神经元的放电,将放电整合到短时窗内,从而组织视觉刺激相关信息。当动物利用V1区视觉信息执行任务时,伽马事件发生率会被选择性调控——这为“根据觉醒状态与行为情境灵活编码感觉信息”提供了框架。因此,围绕离散事件分析神经活动,能为“信号在皮层环路中的精确传递流程”及“这些网络事件的功能影响”提供独特见解。本研究结果也凸显了“基于事件检测皮层活动重复模式”方法的进一步开发潜力:尽管CBASS方法可追踪特定状态下与功率升高相关的事件,但目前该方法仍局限于检测“随时间动态调控的活动模式”。
本研究发现,V1区伽马事件源于对“模式化前馈丘脑-皮层输入”的主动整合[46,47,51]。以往研究普遍认为,伽马等模式化皮层活动由局部兴奋性-抑制性神经元相互作用产生[8,9,21,50,54,55];但皮层加工涉及复杂的局部与长程环路[56],包括丘脑等皮层下结构,以及初级与高级皮层区[10],目前尚不清楚“局部产生的振荡如何在不同加工阶段同步”[29,57,58]。本研究结果提出了一种替代机制:同步化通过“具有特定相位与振幅对齐特征的短时级联同步事件”实现[31]。
总体而言,这些结果为“灵长类[5,32,33,56,57]与啮齿类[25,58,59,60]动物中以往存在争议的研究发现”提供了统一框架,并为“皮层区间功能动态研究”指明了新方向[61,62]。目前,伽马事件发生率与功能影响的复杂调控机制尚未完全明确:我们已证实伽马事件具有丘脑-皮层起源,但V1区其他过程也可能为伽马波段提供能量[63,64];此外,这些网络事件在“不同情境下塑造下游视觉整合与行为反应”中的具体作用,仍需进一步研究。
https://wap.sciencenet.cn/blog-41174-1507928.html
上一篇:首个慢性疲劳综合征血液检测方法问世
下一篇:寿命超过200岁的弓头鲸DNA强修复力的秘密