
氢气通过调节β-羟基丁酸代谢减轻顺铂诱导的肾毒性
背景
肾毒性是许多化疗药物常见的不良反应,也是癌症治疗中剂量限制的主要因素。氢气(H₂)作为一种具有强效抗氧化和抗炎特性的治疗剂应运而生,它能选择性清除羟基自由基并减轻组织损伤。然而,H₂在化疗诱导的急性肾损伤(AKI)中的保护作用及其潜在机制仍知之甚少。
方法
建立顺铂诱导的AKI小鼠模型,分为给予H₂和不给予H₂两组。评估肾损伤生物标志物,并通过TUNEL染色、ELISA和免疫组织化学方法评估炎症和凋亡水平。为探究潜在机制,进行RNA测序,随后通过热图、维恩图和火山图分析识别差异表达基因。利用KEGG通路富集分析探究H₂治疗顺铂诱导的肾毒性时的代谢变化。随后,通过一系列体内和体外实验(包括ELISA、流式细胞术、qRT-PCR、蛋白质印迹和免疫组织化学)验证代谢变化。
结果
吸入H₂通过减少肾组织中的炎症和凋亡,显著减轻顺铂诱导的肾损伤。转录组学分析显示,H₂上调酮体代谢通路,特别是通过增加生酮酶3-羟基-3-甲基戊二酰辅酶A合成酶2(HMGCS2)的表达来增强β-羟基丁酸(β-HOB)的合成。功能实验证实,H₂介导的HMGCS2和β-HOB上调有助于其肾脏保护作用。
结论
氢气通过上调HMGCS2调节β-HOB代谢,从而抑制肾脏炎症和凋亡,对顺铂诱导的肾毒性起到保护作用。这些发现为H₂组织保护作用背后的代谢机制提供了新见解,并为其在减轻化疗诱导的肾损伤中的潜在临床应用提供了理论基础。
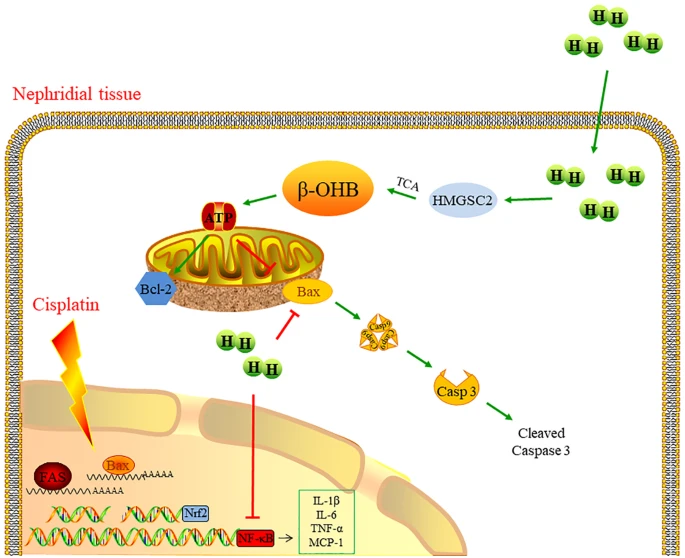
图文摘要
通过顺铂不同剂量和暴露时间处理的肾近端小管细胞中生物标志物HMGB1、NGAL和KIM-1的活性,评估单味中药治疗后肾前性和肾性急性肾损伤的恢复情况
Tian Y, Su H, Chen Y, et al. Molecular hydrogen attenuates cisplatin-induced nephrotoxicity by modulating β-hydroxybutyrate metabolism[J]. Molecular Biology Reports, 2025, 52(1): 1-13.
引言
肾毒性是许多强效化疗药物的严重并发症,约25%-30%的癌症患者会出现顺铂诱导的肾毒性[1,2]。目前,用于预防或减轻肾毒性的有效策略仍然有限。顺铂诱导的氧化应激、细胞衰老和炎症已被认为是肾毒性发展的主要病理事件[1,3]。特别是进行性细胞衰老以及以促炎细胞因子分泌为特征的衰老相关分泌表型(SASP),与多种形式的肾功能障碍密切相关[4]。因此,预防药物诱导的肾损伤是提高化疗药物治疗效果的合理策略。
Ohsawa等人首次报道了氢气(H₂)通过选择性清除羟基自由基而具有神经保护作用,引发了对H₂作为治疗性抗氧化剂的广泛研究[5]。H₂是一种无毒的小分子,在与氧化应激和衰老相关的疾病(包括毒性剂引起的疾病)中具有广泛的治疗潜力[6,7,8]。除了直接的抗氧化特性外,H₂还被证明能减轻炎症并调节细胞死亡信号通路[9]。先前的一项研究表明,吸入H₂气溶胶在脓毒症诱导的急性肾损伤(AKI)模型中表现出抗炎和抗氧化作用,部分是通过抑制肾线粒体分裂[10]。另一项研究报道,H₂通过激活核因子 erythroid 2相关因子2(Nrf2)通路发挥抗衰老作用[11]。此外,氢气和富氢水已被证明能减轻顺铂诱导的肾毒性,且不影响其抗肿瘤 efficacy[12,13]。然而,这些保护作用背后的详细分子机制仍不完全清楚。值得注意的是,线粒体是细胞能量代谢的中心枢纽[14]。在厌氧条件下,酮体主要在肝脏和肾脏中合成,在维持线粒体能量稳态和膜电位稳定性方面起着至关重要的作用[15,16,17]。H₂的保护作用是否涉及调节酮体合成的线粒体通路仍是一个悬而未决的问题。
在本研究中,我们进一步探讨了H₂在顺铂诱导的小鼠肾毒性模型中的潜在保护作用。我们发现H₂通过上调3-羟基-3-甲基戊二酰辅酶A合成酶2(HMGCS2)在调节β-羟基丁酸(β-HOB)代谢中发挥关键作用。我们的发现为H₂介导的肾脏保护作用提供了机制见解,并为其作为减轻化疗诱导的肾损伤的安全有效策略的潜在应用提供了实验证据。
材料和方法
细胞与实验设计
人近端小管细胞(HK-2)购自上海世宏生物科技有限公司。细胞在添加10%胎牛血清(FBS;Gibco,美国)和1%青霉素-链霉素溶液(Gibco,美国)的Dulbecco改良Eagle培养基(DMEM)(Gibco,美国)中培养。处理时,将细胞置于含有不同浓度H₂(10%、20%或30%)的混合气体(H₂、21% O₂和5% CO₂)培养箱中。以0% H₂、21% O₂和5% CO₂的混合气体作为对照。
细胞活力测定
将细胞以每孔6000个的密度接种在96孔板中,孵育24小时。随后,用H₂(0、10%、20%、30%)预处理24小时,然后暴露于20μM顺铂或不暴露。孵育指定时间后,应用CCK-8试剂盒(TargetMol,美国),在450nm处检测吸光度值。
细胞凋亡测定
用0或30% H₂预处理24小时后,HK-2细胞用20μM顺铂处理。孵育6小时后,将细胞收集在结合缓冲液中,使用Annexin V-FITC/PI凋亡检测试剂盒(上海世宏生物,中国)用5μL碘化丙啶(PI)和Annexin V-Alexa Fluor 488标记。通过流式细胞术(Beckman,美国)测定凋亡细胞的比例。
动物与实验设计
体内研究使用C57BL/6雄性小鼠,平均体重20g。所有操作均遵循《实验室动物护理原则》,并经同济大学医学院附属上海第十人民医院动物研究委员会批准(中国上海)。小鼠随机分为四个实验组(每组n=8):[1]对照组:小鼠在第10天腹腔内(i.p.)注射无菌生理盐水;[2]H₂组:小鼠连续13天每天暴露于30% H₂(平衡空气/氧气)中6小时;[3]顺铂组:小鼠在第10天腹腔内单次注射顺铂(20mg/kg,江苏豪森药业股份有限公司);[4]H₂+顺铂组:小鼠在第0天用30% H₂预处理(每天6小时)。在吸入期间的第10天给予顺铂(20mg/kg,腹腔注射)[20]。实验设计示意图见图S1A。使用AMS-H-01氢氧雾化器(Asclepius,上海,中国),通过COSMOS XP-3140检测器(日本)连续监测浓度并维持在30%。为进一步研究H₂的肾脏保护机制,我们设置了对照组、顺铂组和β-HOB+顺铂组。其中,小鼠在第10天给予顺铂前,连续12天用β-HOB(20mmol/kg,腹腔注射)预处理[21]。实验设计示意图见图S1B。
肾功能评估
从小鼠眼眶静脉丛采集血液样本,离心(3000rpm,15分钟,4°C)分离。使用市售检测试剂盒(南京建成,中国)按照制造商的方案测定血尿素氮(BUN)和肌酐(Cr)水平。
组织病理学检查
收集肾脏组织并进行苏木精-伊红(HE)染色。根据肾小管细胞空泡变性和脱落程度评估损伤,评分标准如下:无损伤=0;<25%=1;<50%=2;<75%=3;>75%=4。
末端脱氧核苷酸转移酶dUTP缺口末端标记(TUNEL)染色
使用TUNEL检测试剂盒(Beyotime,上海,中国)检测凋亡细胞。将细胞与TUNEL反应混合物孵育,随后用DAPI复染并拍照。
炎症细胞因子测定
使用相关ELISA试剂盒(CUSABIO,武汉,中国)分析白细胞介素-1β(IL-1β)、白细胞介素-6(IL-6)、单核细胞趋化蛋白-1(MCP-1)和肿瘤坏死因子-α(TNF-α)的蛋白水平。通过离心收集单细胞悬液,然后按照相关方案进行操作。使用酶标仪(Molecular Devices,美国)在450nm处测量吸光度。
蛋白质印迹分析
收集肾脏组织,按照制造商的方案进行蛋白质印迹分析。靶向HMGCS2的一抗(Cell Signaling Technology,美国)和HRP偶联的二抗稀释至1:1000浓度。以β-肌动蛋白(Cell Signaling Technology,美国)作为内参进行蛋白质印迹分析。
RNA测序分析
分析对照组、顺铂组和H₂处理组肾脏组织的差异表达基因(DEGs)。测序数据使用Illumina平台生成。 fold change(FC)≥2且错误发现率(FDR)<0.05的基因被认为是显著差异表达基因。使用京都基因与基因组百科全书(KEGG)数据库进行通路和基因本体论(GO)分析。
RNA提取、cDNA合成和实时PCR定量
步骤如先前报道[22]。从肾脏组织样本中提取RNA,并逆转录为cDNA。PCR扩增和测序由上海美吉生物医药科技有限公司使用Illumina Novaseq 6000完成。引物序列见表1。采用比较Ct(ΔΔCT)法分析数据。
表1 实时PCR分析所用引物
统计分析
数据以均数±标准差(SD)表示。组间差异采用单因素方差分析(ANOVA)检验。P值<0.05被认为具有统计学意义。使用GraphPad Prism 7软件进行统计分析。
结果
氢气减轻顺铂诱导的肾毒性
为评估氢气对顺铂诱导的肾毒性的保护作用,首先对肾脏组织进行了组织病理学分析。正如预期的那样,顺铂处理小鼠的HE染色切片显示出广泛的肾小管损伤,包括上皮细胞坏死、刷状缘丢失、肾小管扩张和炎症细胞浸润(图1A)。经氢气预处理的小鼠,其这些病理特征得到显著改善,表现为结构损伤减轻和肾小管损伤评分降低(图1B)。同时,检测了肾功能的生化标志物。顺铂给药导致血尿素氮(BUN)和血清肌酐(Cr)水平显著升高,而氢气预处理显著减轻了这种升高(图1C和D)。总之,这些结果表明氢气对顺铂诱导的肾毒性具有显著的保护作用。
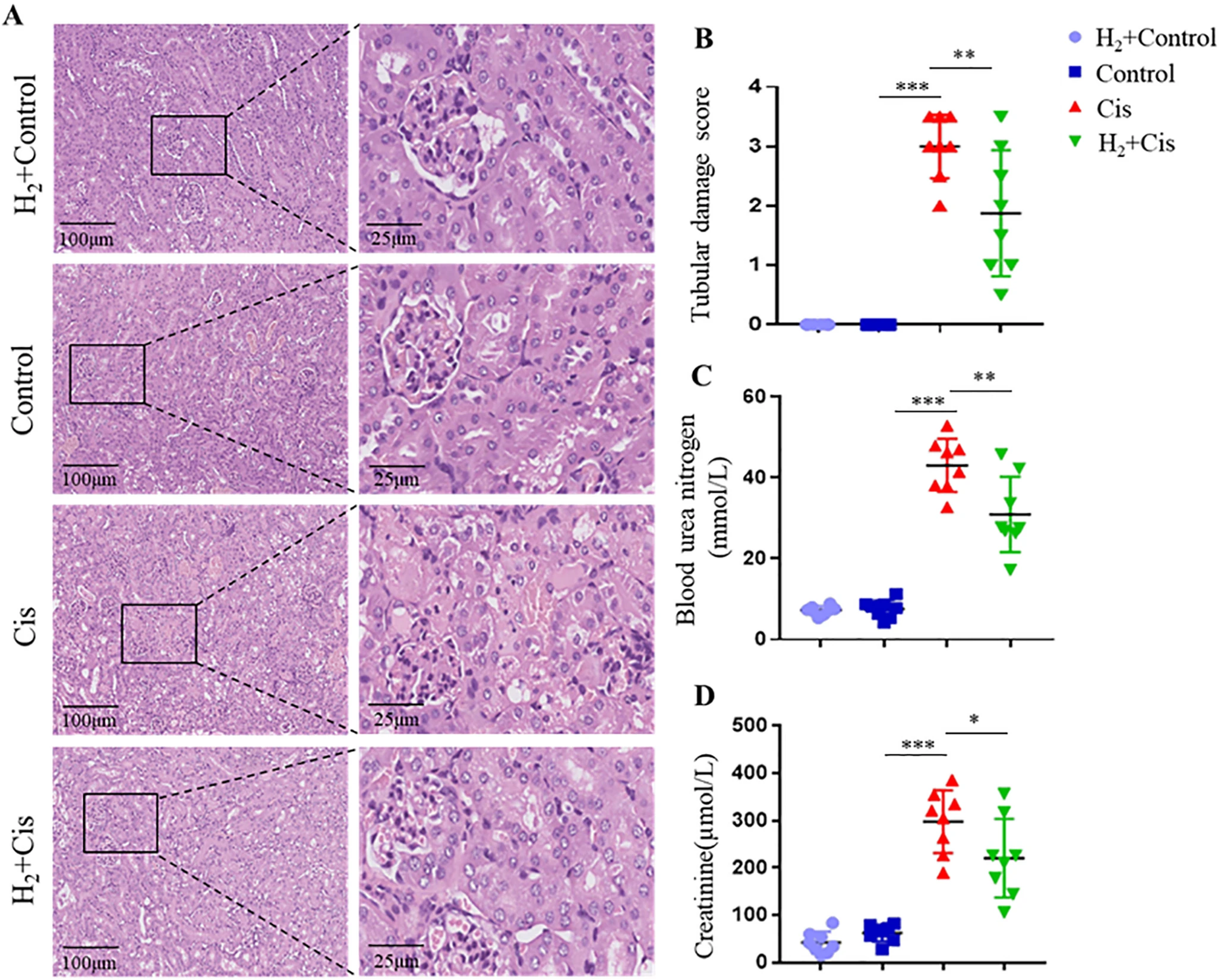
图1 氢气减轻小鼠顺铂诱导的肾毒性。(A)肾脏切片的代表性HE染色,显示组织病理学变化。(B)肾小管损伤评分的定量分析。(C和D)不同处理组的血清BUN(C)和Cr(D)水平。*p < 0.05,**p < 0.01,***p < 0.001。缩写:BUN,血尿素氮;Cr,肌酐;Cis,顺铂
气雾化氢气减轻顺铂诱导的炎症和凋亡
为确定氢气如何减轻顺铂诱导的肾损伤,我们检测了肾脏组织中促炎细胞因子(如IL-1β、IL-6、MCP-1和TNF-α)的水平。ELISA检测显示,顺铂处理后这些促炎介质的水平显著升高(图2A-D)。吸入氢气显著抑制了这些细胞因子的水平,表明其具有强效的抗炎作用(图2A-D)。鉴于凋亡是顺铂诱导的肾毒性的另一个关键病理特征,我们在顺铂注射后72小时使用TUNEL染色进一步评估了凋亡细胞死亡情况。顺铂组中观察到TUNEL阳性细胞显著增加,而氢气给药显著减少了这种增加(图2E和F)。这些发现共同表明,气雾化氢气通过同时抑制炎症和凋亡来减轻顺铂诱导的肾损伤。
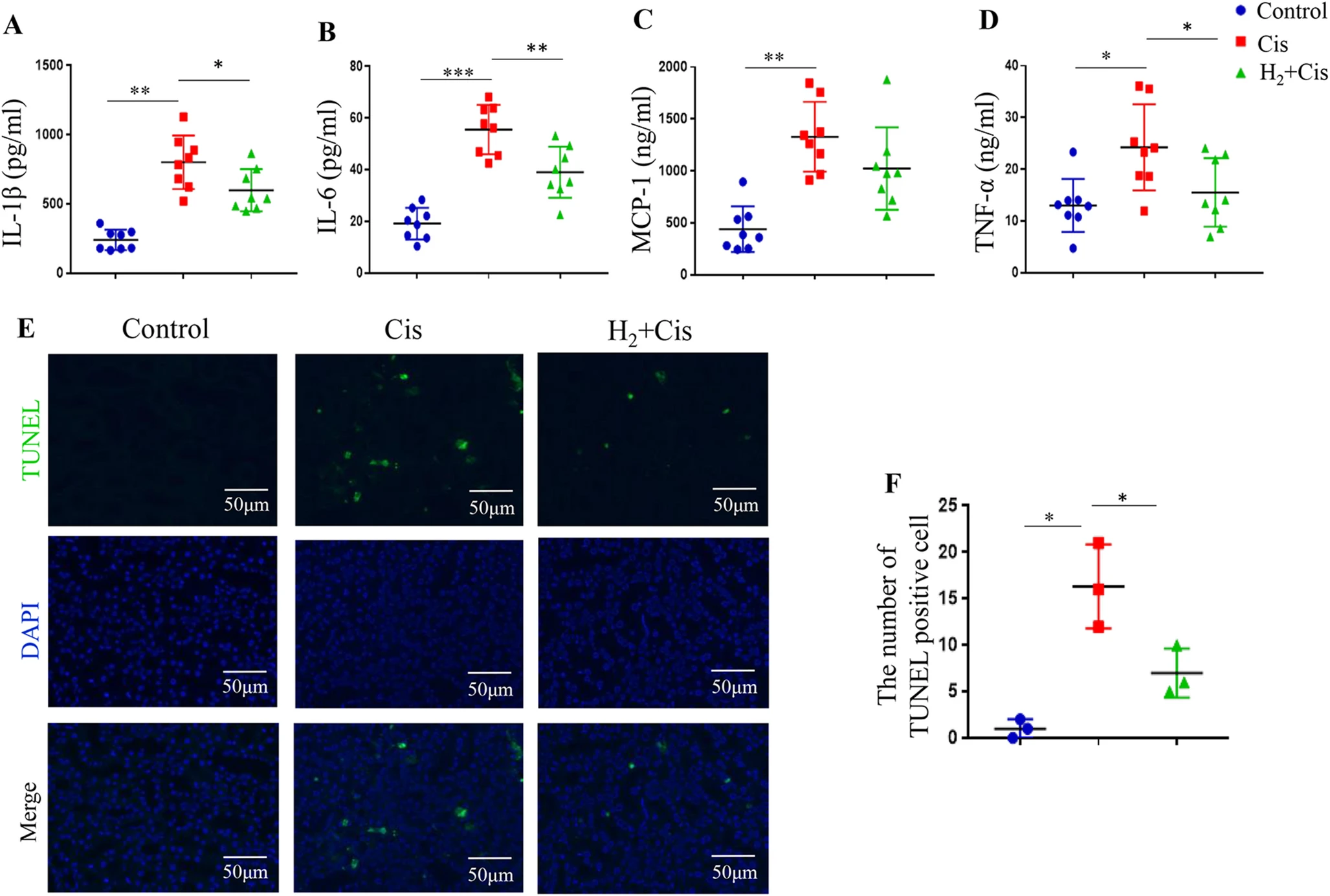
图2 气雾化氢气给药减少顺铂诱导的炎症和凋亡。(A-D)ELISA检测肾脏组织中炎症细胞因子IL-1β(A)、IL-6(B)、MCP-1(C)和TNF-α(D)的水平。(E)显示凋亡细胞的代表性TUNEL染色图像。(F)TUNEL阳性细胞的定量分析。*p < 0.05,**p < 0.01,***p < 0.001。缩写:Cis,顺铂;TUNEL,末端脱氧核苷酸转移酶dUTP缺口末端标记
氢气处理顺铂诱导的肾毒性中的差异基因表达分析
为探究氢气介导的保护作用的潜在分子机制,对肾脏组织进行了RNA测序转录组分析。如热图和维恩图所示,实验组之间观察到不同的基因表达模式(图3A和B)。火山图显示,与单独接受顺铂的组相比,氢气处理的顺铂组中有472个显著上调的基因和556个显著下调的基因(图3C)。通过qRT-PCR测定验证了代表性基因,包括上调的Lce1d和Il1rn以及下调的Hbb-bs和Akr1c21(图3D-G),证实了转录组分析对于后续通路探索的可靠性。
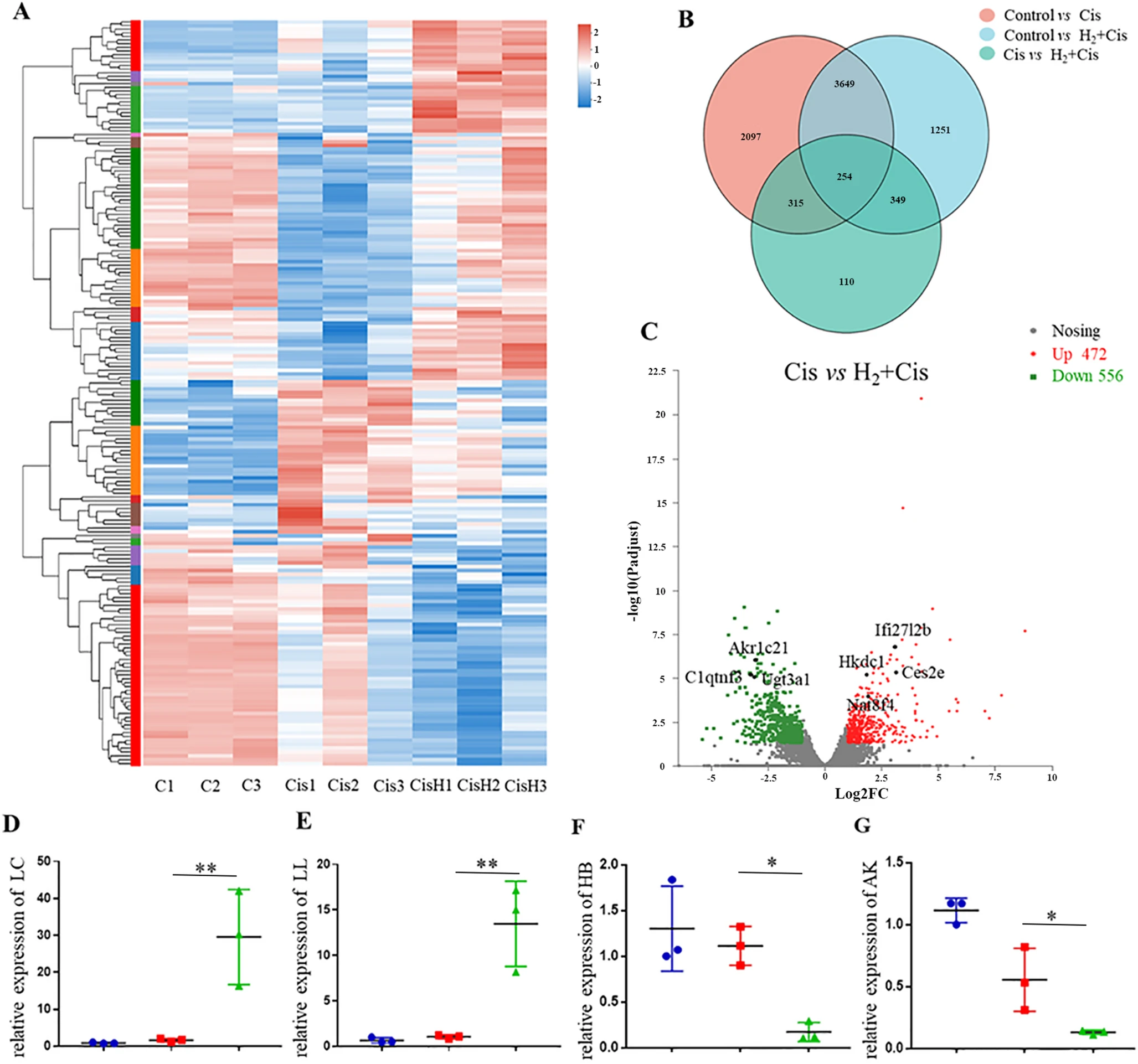
图3 转录组分析揭示氢气处理顺铂诱导的肾毒性时的差异基因表达。(A)实验组间差异表达基因(DEGs)的热图。(B)显示共有和独特差异表达基因的维恩图。(C)火山图显示顺铂组和氢气+顺铂组之间显著上调和下调的基因。(D-G)所选差异表达基因的qRT-PCR验证:Lce1d(D)、Il1rn(E)、Hbb-bs(F)和Akr1c21(G)。*p < 0.05,**p < 0.01。缩写:DEGs,差异表达基因
β-羟基丁酸代谢参与体内氢气介导的肾脏保护作用
为阐明顺铂组和氢气+顺铂组之间1028个差异表达基因相关的生物学功能和通路,使用DAVID数据库进行了基因本体论(GO)和京都基因与基因组百科全书(KEGG)通路富集分析。GO术语显示,分子功能中最显著富集的术语主要分别与结合活性、细胞成分和细胞过程相关(图4A)。此外,KEGG通路分析确定了20个最显著富集的通路。其中,在顺铂处理的小鼠中,包括酮体代谢、丁酸代谢和三羧酸(TCA)循环在内的代谢通路显著富集,表明β-羟基丁酸代谢可能参与其中(图4B)。随后的ELISA测定证实,氢气给药后β-羟基丁酸水平显著升高(图4C)。同时,作为β-羟基丁酸代谢中的关键辅因子,烟酰胺腺嘌呤二核苷酸(NAD⁺)的水平也显著增加(图4D),而乙酰辅酶A(acetyl-CoA)水平呈上升趋势,但氢气给药组未达到统计学显著性(图4E)。为进一步验证β-羟基丁酸的功能作用,向顺铂处理的小鼠施用外源性β-羟基丁酸。值得注意的是,补充β-羟基丁酸显著减轻了肾损伤,与氢气的保护作用相似(图4F和G)。这些发现强调了β-羟基丁酸代谢在氢气介导的肾脏保护中的关键作用。

图4 β-羟基丁酸代谢参与体内氢气的肾脏保护作用。(A)使用RNA测序进行的靶基因GO分类。(B)20个最显著富集的KEGG通路分类。(C-E)肾脏组织中β-羟基丁酸(C)、NAD⁺(D)和乙酰辅酶A(E)水平的ELISA定量。(F)对照组、顺铂组和β-羟基丁酸+顺铂组肾脏切片的代表性组织学图像。(G)肾小管损伤评分的定量分析。*p < 0.05,**p < 0.01,***p < 0.001。缩写:Cis,顺铂;β-HOB,β-羟基丁酸;GO,基因本体论;KEGG,京都基因与基因组百科全书;NAD⁺,烟酰胺腺嘌呤二核苷酸;A-CoA,乙酰辅酶A
β-羟基丁酸代谢有助于体外氢气介导的细胞保护作用
为进一步研究β-羟基丁酸代谢的作用,建立了使用人近端小管细胞(HK-2)的体外模型。20μM顺铂处理显著抑制HK-2细胞增殖,而与氢气共同处理以剂量依赖方式减轻这种细胞毒性作用(图5A)。使用膜联蛋白V/碘化丙啶(Annexin V/PI)试剂盒的流式细胞术分析证实,30%氢气显著减少顺铂处理细胞的凋亡(图5B和C)。在细胞形态方面,30%氢气+对照组和对照组为梭形,这是典型的上皮样细胞形态。而顺铂处理的细胞呈现圆形和皱缩,30%氢气共同处理明显缓解了这些特征(图5D)。与体内发现一致,30%氢气处理后细胞内β-羟基丁酸和NAD⁺水平显著增加(图5E-G),进一步证实了β-羟基丁酸代谢在氢气介导的细胞保护中的作用(图5E-G)。
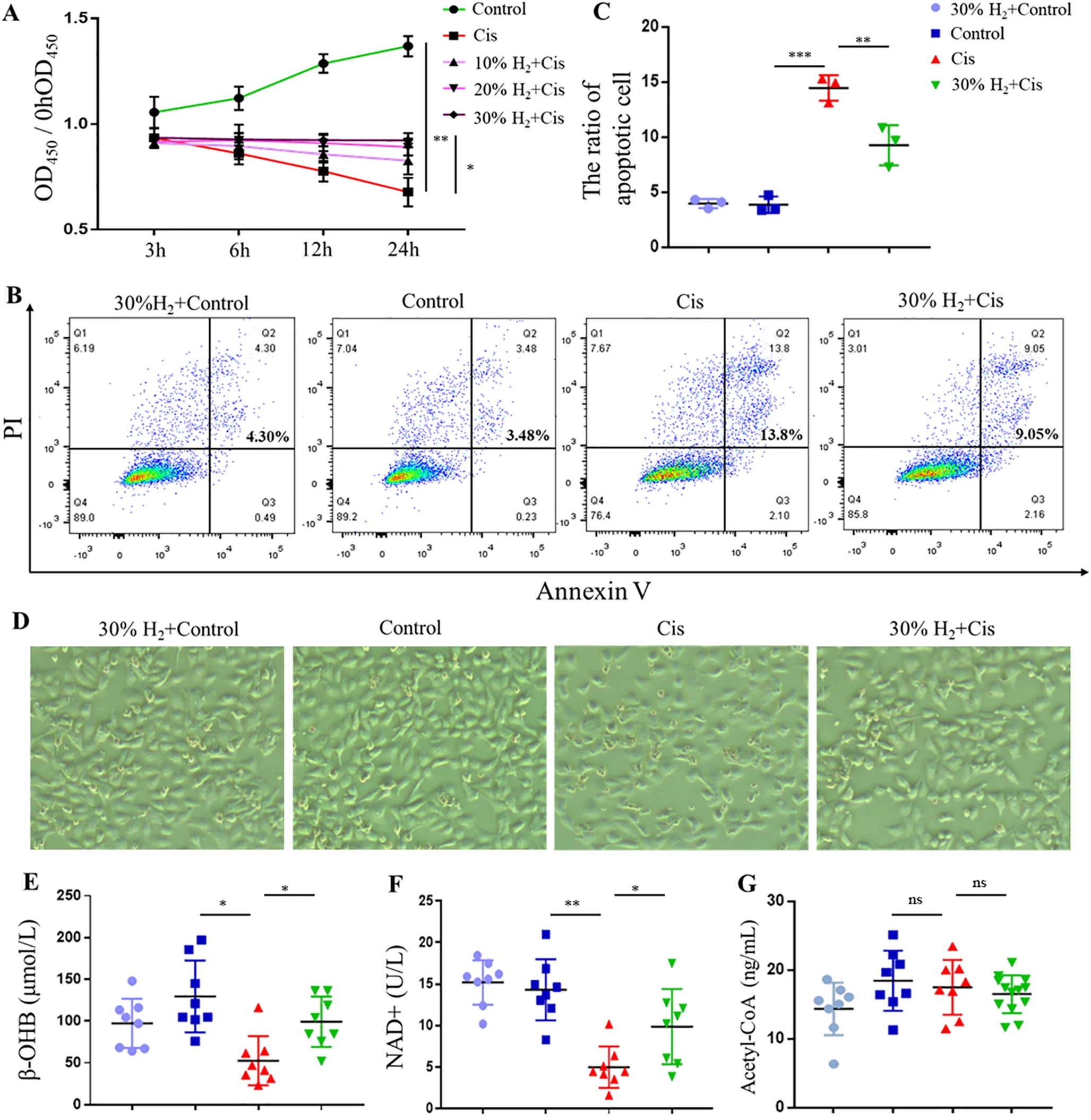
图5 β-羟基丁酸代谢有助于氢气对HK-2细胞的细胞保护作用。(A)不同氢气浓度下通过CCK-8测定的细胞活力。(B)使用碘化丙啶/膜联蛋白V染色的流式细胞术凋亡分析。(C)凋亡细胞的定量分析。(D)每组的代表性细胞形态。(E-G)细胞内β-羟基丁酸(E)、NAD⁺(F)和乙酰辅酶A(G)的ELISA测定。*p < 0.05,**p < 0.01。缩写:β-HOB,β-羟基丁酸;NAD⁺,烟酰胺腺嘌呤二核苷酸;A-CoA,乙酰辅酶A
氢气上调3-羟基-3-甲基戊二酰辅酶A合成酶2以增强酮体代谢
为阐明氢气调节β-羟基丁酸代谢的机制,我们检测了生酮作用中的限速酶3-羟基-3-甲基戊二酰辅酶A合成酶2(HMGCS2)的表达。免疫组织化学染色显示,顺铂损伤的肾组织中HMGCS2显著下调,而氢气给药后显著恢复(图6A和B)。在HK-2细胞中,顺铂抑制HMGCS2的蛋白质和mRNA水平,但氢气给药后显著上调(图6C-E)。为进一步验证HMGCS2的功能重要性,在HK-2细胞中设置对照组+ hymeglusin(HMGCS2抑制剂)组(用20μM hymeglusin处理4小时);对照组+ hymeglusin +氢气组(用20μM hymeglusin处理4小时,然后用30%氢气处理6小时)。蛋白质印迹测定显示,在存在hymeglusin的情况下,氢气处理可显著提高HMGCS2的水平和基因表达(图6F-H)。总之,这些发现表明氢气通过增强酮体代谢,特别是通过β-羟基丁酸生成和HMGCS2调节,来保护免受顺铂诱导的肾毒性。
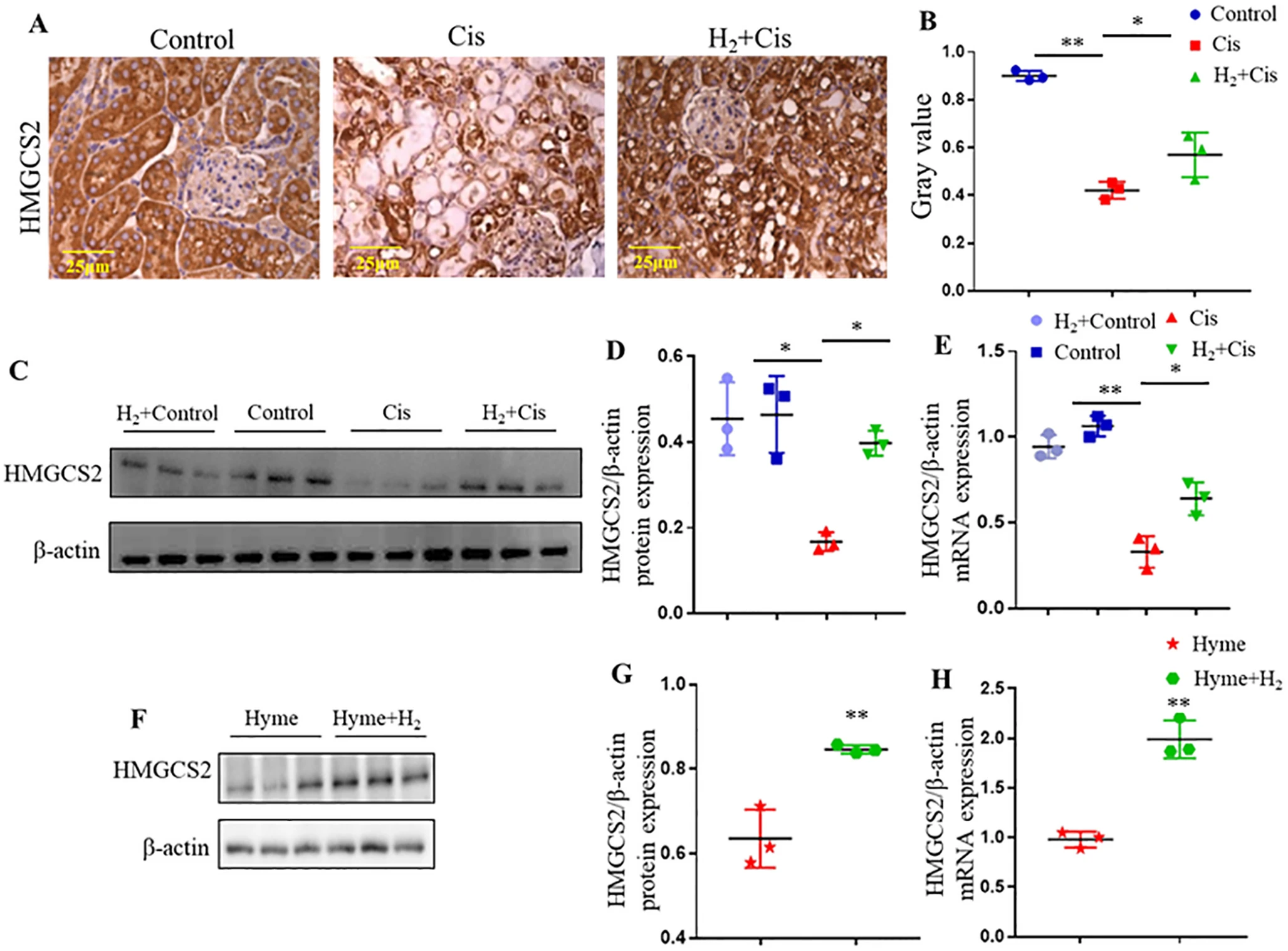
图6 氢气在体内和体外促进HMGCS2表达。(A)肾脏组织中HMGCS2的免疫组织化学染色。(B)HMGCS2免疫反应性的定量分析。(C)HK-2细胞中HMGCS2蛋白的蛋白质印迹分析。(D)蛋白质印迹结果的密度分析。(E)通过qRT-PCR测定的HK-2细胞中HMGCS2的mRNA表达。(F-H)用hymeglusin(HMGCS2抑制剂)±氢气处理的HK-2细胞中HMGCS2的蛋白质(F和G)和基因(H)表达。*p < 0.05,**p < 0.01。缩写:Cis,顺铂;HMGCS2,3-羟基-3-甲基戊二酰辅酶A合成酶2;Hyme,hymeglusin
讨论
在本研究中,我们证明氢气在体内和体外均对顺铂诱导的肾毒性具有显著的保护作用。氢气预处理显著减轻肾脏组织病理学损伤,降低血清BUN和肌酐水平,并抑制炎症细胞因子表达和凋亡。从机制上讲,氢气处理上调HMGCS2表达并增加β-羟基丁酸和NAD⁺水平,从而将能量代谢转向生酮作用,并抑制线粒体氧化应激和炎症。
顺铂是一种用于治疗各种实体瘤的强效化疗药物,但其临床疗效因剂量依赖性肾毒性而显著受限[23]。临床上,顺铂诱导的肾毒性的特征是肾清除率降低、低镁血症和低钾血症[24]。从机制上讲,Amini团队进一步揭示,顺铂通过诱导炎症、氧化应激和凋亡,同时抑制Sirtuin1/Nrf2/HO-1通路来促进肾功能障碍[25,26]。一致地,我们的发现也证实了顺铂诱导的肾损伤,组织学上的肾小管损伤以及BUN和血清肌酐水平升高均可证明这一点。与传统抗氧化剂相比,氢气具有几个独特的优势。作为一种小的中性分子,氢气可以快速扩散穿过细胞膜并分布到整个亚细胞区室[27]。氢气已被证明在各种炎症和氧化应激相关疾病中具有保护作用,包括急性腹膜炎[28]和创伤性脑损伤[29],主要通过减轻核因子κB(NF-κB)介导的炎症和调节凋亡通路,涉及DNA损伤和Bax/半胱天冬酶(caspase)激活[30,31,32]。然而,仅这些抗氧化作用可能无法完全解释氢气广泛的保护作用。最近的研究表明,氢气还可能影响线粒体能量代谢[33]。线粒体不仅是细胞内活性氧(ROS)的主要来源,也是三磷酸腺苷(ATP)生产和代谢调节的中心枢纽[34]。几项研究表明,氢气可以通过激活线粒体融合蛋白-2(Mfn2)和过氧化物酶体增殖物激活受体γ辅激活因子-1α(PGC-1α)等通路来维持线粒体膜电位,促进ATP生成,并改善线粒体动力学和生物发生[35]。在心肌组织中,氢气上调氧化磷酸化中的关键电子载体辅酶Q9(CoQ9),增强ATP合成并保护免受缺血性损伤[36]。我们的研究证实,吸入氢气显著改善顺铂诱导的组织学损伤、升高的BUN和肌酐水平、肾脏炎症和肾小管凋亡。这些发现与先前关于氢气对急性肾损伤的保护作用的报道一致[12,13]。值得注意的是,细胞衰老已成为肾损伤的关键病理机制,其特征是衰老相关分泌表型(SASP)因子的分泌,这会导致组织功能障碍和纤维化重塑。我们的研究还表明,氢气降低了SASP因子(IL-1β、IL-6、TNF-α)的水平[37]。同样,原等人也证明氢气对血管内皮细胞具有持久的抗氧化和抗衰老作用[38]。氢气减轻凋亡和衰老通路的能力表明其具有比以前所认识到的更广泛的细胞保护作用。在这种情况下,我们的发现揭示了氢气在顺铂诱导的肾毒性模型中的保护作用。通过RNA测序分析,我们在氢气处理的肾脏中发现了独特的转录组特征,其中参与酮体代谢和线粒体功能的基因显著富集。其中,生酮作用中的限速酶HMGCS2的上调尤为值得注意。
酮体(β-羟基丁酸、乙酰乙酸和丙酮)是在碳水化合物摄入有限或糖异生氨基酸可用性降低时由肝脏大量合成的生理代谢物[39]。除肝脏外,肾脏也可能生成酮体[39]。除了在营养缺乏时作为替代能量底物外,酮体还作为信号代谢物调节基因表达、炎症和细胞应激反应[40]。β-羟基丁酸是主要的酮体,约占循环中酮体的80%[41]。在用β-羟基丁酸处理的老年大鼠中,促炎信号被逆转,同时叉头框蛋白O1(FoxO1)及其下游靶标过氧化氢酶和锰超氧化物歧化酶(MnSOD)上调,而PGC-1α选择性地与FoxO1相互作用以减轻肾脏炎症[42]。此外,有证据表明,升高的血清β-羟基丁酸降低人单核细胞或小鼠炎症模型中NLRP3炎症小体介导的半胱天冬酶-1激活以及IL-1β和IL-18的产生[43]。最近的研究发现,低浓度的内源性β-羟基丁酸补充通过改变线粒体ATP代谢来缓解氧化应激,从而减轻小鼠肾损伤[44]。我们的发现支持并扩展了这一概念,表明氢气通过诱导HMGCS2升高内源性β-羟基丁酸水平,从而将能量代谢转向生酮作用,并抑制线粒体氧化应激和炎症。科学证据还支持使用外源性酮体和其他形式的酮补充剂,作为限制性饮食方案的辅助手段或作为生酮饮食的潜在替代方案。这些代谢调整伴随着NAD⁺水平的增加,NAD⁺是线粒体代谢和去乙酰化酶激活中的重要辅因子,进一步表明生物能量能力增强。此外,已证实N-乙酰半胱氨酸(NAC)通过抑制促炎C5a受体,在顺铂诱导的急性肾损伤小鼠中发挥抗氧化和抗炎作用[45]。出乎意料的是,作为β-羟基丁酸代谢的下游产物,乙酰辅酶A水平没有显著变化。这可能是由于酮代谢的组织特异性差异,因为肝脏是β-羟基丁酸合成的主要部位,而肾脏主要利用β-羟基丁酸作为底物。氢气可能促进肝脏生酮作用,并通过循环促进β-羟基丁酸向肾组织的运输,这一假设值得进一步研究。
结论
我们的研究表明,氢气在体内和体外均对顺铂诱导的肾毒性具有显著的保护作用。从机制上讲,氢气降低促炎细胞因子水平并减轻凋亡。转录组分析确定了差异表达基因和富集的代谢通路,尤其涉及酮体代谢。其中,氢气给药后β-羟基丁酸水平显著升高。重要的是,顺铂下调HMGCS2的表达,而氢气处理显著恢复其表达,表明HMGCS2介导的β-羟基丁酸代谢在氢气的肾脏保护机制中起关键作用。此外,我们认为外源性酮补充或饮食干预可能作为氢气治疗在临床环境中的补充策略。
转载本文请联系原作者获取授权,同时请注明本文来自孙学军科学网博客。
链接地址:https://wap.sciencenet.cn/blog-41174-1495462.html?mobile=1
收藏