博文
内质网-线粒体接触位点研究取得进展
||
内质网-线粒体接触位点研究取得进展
内质网线粒体接触点是位于内质网膜与线粒体外膜之间通过蛋白质介导的物理性接触部位。参与脂质合成、钙离子信号、线粒体生物发生、细胞凋亡及胞内转运等重要细胞生理过程的调节。
细胞内各种细胞器,如内质网和线粒体之间的快速通讯对于细胞的协调功能至关重要。这种相互作用主要发生在隔间直接接触的部位,并由特定的系留机制介导。高速单分子跟踪揭示了一个调节良好的界面,在该界面上,许多快速结合和解结合事件提供了高度适应性的相互作用。
摘要:为了协调细胞生理学,真核细胞依赖于特定细胞器-细胞器接触位点的分子快速交换1,2.内质网-线粒体接触位点 (ERMCS) 是特别重要的通信枢纽,在信号分子、脂质和代谢物的交换中发挥着关键作用3,4.ERMCS通过每个细胞器表面互补的系系分子之间的相互作用来维持5,6.然而,由于这些膜界面对实验扰动极敏感7,8,仍然缺乏对其纳米级组织和调控的清晰认识。在这里,我们将三维电子显微镜与模型细胞器系绳、囊泡相关膜蛋白 (VAMP) 相关蛋白 B (VAPB) 的高速分子跟踪相结合,以绘制 ERMCS 的结构和扩散图。我们在VAPB接触位点中发现了与内质网膜曲率相关并经历快速重塑的动态子域。我们发现,VAPB分子在几秒钟内进入和离开ERMCS,尽管接触位点本身在更长的时间尺度上保持稳定。这种亚稳态使ERMCS能够随着生理环境的变化而重塑,以适应细胞的代谢需求。VAPB 中肌萎缩侧索硬化症相关的突变会扰乱这些亚结构域,可能损害它们的重塑能力并导致细胞器间通讯受损。这些结果将高速单分子成像确立为绘制接触位点界面结构的新工具,并揭示了VAPB在接触位点的扩散景观是ERMCS稳态的关键组成部分。
问题提出
真核生物中膜封闭的细胞器 - 具有具有细胞核的生物体 - 需要特定的机制来交流和同步在物理分离的隔室中发生的生化反应。这种通讯主要发生在细胞器彼此直接接触的部位1.在这些位点,细胞器彼此靠近但不融合,它们之间的相互作用由分子系介导,分子系提供蛋白质-蛋白质或蛋白质-脂质连接。蛋白质合成机制 - 内质网(ER) - 和称为线粒体的能量产生细胞器之间的接触至关重要,因为这些界面协调基本的代谢和信号传导功能。如何调节介导这些接触的系绳2仍然不清楚,主要是因为这些接触位点小于衍射极限(通过光学显微镜可以区分两个物体的最小距离),并且许多系绳在不同细胞器的接触位点之间快速移动。
解决方案
我们开发了一种改进版的单颗粒跟踪光活化定位显微镜3,4.我们同时跟踪单个系系分子,并收集有关ER和线粒体在同一细胞中的位置的信息。通过将单个内质网包埋蛋白的轨迹映射到内质网和线粒体结构上,我们能够直接观察和识别细胞器接触位点的拴系相互作用。
我们使用了一种高度保守且表征良好的内质网拴系蛋白,称为VAPB及其线粒体结合伙伴PTPIP51,作为模型系统。我们发现VAPB介导的拴系相互作用特别依赖于这两个分子的结合(图1a)。
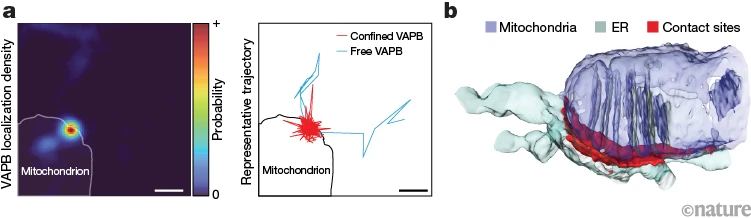
图 1 |许多动态的单分子相互作用共同产生了内质网和线粒体之间接触位点的结构。 a,左图,内质网 (ER) 和线粒体之间接触位点的 VAPB 定位密度,使用一分钟内的许多单独轨迹计算。右图是单个 ER 定位的 VAPB 分子的轨迹,显示了 VAPB 与线粒体的结合位置(红色迹线)以及它在 ER 表面上自由扩散的位置(蓝色迹线)。比例尺,500纳米。b,使用具有代表性的内质网-线粒体接触位点的电子显微镜进行 3D 重建。图片来源:Obara, C. J. et al./自然 (CC BY 4.0的)
这些相互作用揭示了接触位点内复杂的扩散景观,这与3D电子显微镜重建中细胞器的形状直接相关(图1b)。VAPB介导的内质网-线粒体接触位点通过许多高度瞬态的相互作用动态组装 - 大多数分子在接触位置停留不到600毫秒 - 这些相互作用共同产生粘附力,使两个膜保持数十秒。由此产生的界面具有高度的移动性,可以在几秒钟的时间尺度上轻松适应细胞需求的变化。事实上,我们发现VAPB拴系机制中与疾病相关的突变降低了这些界面上系绳的交换速率,可能会扰乱细胞信号在位点的调节效率。
影响
这些发现有助于展示细胞器的接触位点如何被精确调节,同时仍然能够在几秒钟内重新排列。由于接触位点的系系与细胞器其余部分的未结合池交换得如此迅速,因此预计对未结合系系蛋白的生化修饰将很快包含在结合群体中,从而导致接触的物理性质发生变化。这种动态反应似乎对细胞对代谢信号的快速反应很重要。我们的观察还强调了充分了解分子行为的重要性。例如,尽管大多数与疾病相关的突变VAPB分子存在于胞质溶胶的聚集体中,但其中一些分子浸润了接触位点,这表明病理学可能由这些蛋白质的一小部分驱动。
本研究重点关注了最具特征的内质网-线粒体拴系对。未来的研究将需要检查许多其他已知系绳的作用,包括内质网和线粒体之间,以及不同组织和环境中的其他细胞器接触位点。使用这种方法直接观察这些相互作用是如何被调节的也将是令人兴奋的,因为这种调节对所有真核细胞的代谢功能至关重要。这种知识在疾病方面尤为重要,因为许多来源不明的遗传疾病可能是由生物机制的信号或动力学变化引起的,而这些变化可能仅在单分子水平上可见。— Christopher J. Obara 和 Jennifer Lippincott-Schwartz 在美国弗吉尼亚州阿什本的霍华德休斯医学研究所。
专家意见
这项巨大的工作,将单颗粒跟踪与体积成像相结合,使用聚焦离子束扫描电子显微镜来关联结构和功能,非常新颖和令人兴奋。此外,对方法的仔细和高度详细的描述,特别是对补充信息中陷阱的讨论,使这份手稿成为一项重要贡献,将对其他研究细胞器-细胞器接触的研究人员产生严重影响。——莎伦·沃尔夫(Sharon G. Wolf)在以色列雷霍沃特的魏茨曼科学研究所(Weizmann Institute of Science)工作。
论文背后故事
这项工作是多机构协作重要性的一个很好的例子。我们开始了一个“大胆创新计划”项目来了解这个系统,这个系统很难分析——这远远超出了任何一个团队的能力。只有通过许多具有不同技能的人的贡献,包括光学显微镜、细胞生物学、生物物理学、计算建模和分析以及电子显微镜,我们才能够理解这个系统。特别是,我们很幸运地得到了乔纳森·尼克松-阿贝尔(Jonathon Nixon-Abell)和克雷格·布莱克斯通(Craig Blackstone)的指导,他们参与了每一步,并有远见地意识到这种方法可能与人类疾病直接相关。在未来的几年里,我们将看到将个体分子行为与疾病病理学联系起来是否有助于理解疾病机制,这将是令人惊讶的。— C.J.O. 和 J.L.-S.
编者寄语
细胞器间接触位点既动态又小,这使得其详细的功能分析具有挑战性。因此,特别令人兴奋的是,Obara及其同事提供了对最热门的接触部位之一(ER和线粒体之间的接触部位)的一般行为,定位和压力反应的复杂见解。— Sadaf Shadan,《自然》杂志高级编辑兼团队经理
部分正文:
哺乳动物细胞中细胞器之间最普遍的接触部位是内质网 (ER) 和线粒体之间9,10.这种界面与健康和疾病中的多种生物过程有关,从脂质合成和分解代谢到钙信号传导和促进细胞呼吸2、3、11.这些结构通常被称为内质网-线粒体接触位点(ERMCSs)或线粒体-内质网接触位点(MERC),在真核生物中无处不在,可能以几种不同的形式存在(完整讨论见补充文本,第2a-b节)3、4、6、12.然而,技术局限性限制了我们对ERMCS生物学许多方面的理解,因为其结构对固定伪影非常敏感,即使在单个细胞中也显示出显着的生物学异质性。因此,基于生物化学的方法经常受到平均伪影的影响,而且大多数现有的标签技术都会干扰ERMCS的大小、结构和监管(见补充文本,第1节和第2节的讨论)7.特别是,我们缺乏对其分子成分和亚结构的动态调控的理解。在这里,我们介绍了结合三维(3D)电子显微镜和活细胞高速单分子成像的定量成像方法,以描述ERMCS内分子在不同生理和病理条件下的亚结构和动力学。
内质网和线粒体之间的特定接触位点在反式中由成对的分子链介导(图1)。1一个)1,5,6.一种成熟的 ERMCS 系绳是 ER 定位的囊泡相关膜蛋白 (VAMP) 相关蛋白 B (VAPB)13、14、15 ,与线粒体结合伙伴相互作用,促进钙和脂质转移16,17并且可能携带导致严重运动神经元疾病的突变18,19.VAPB在线粒体中的最佳结合伙伴是线粒体外膜(OMM)成分,即蛋白酪氨酸磷酸酶相互作用蛋白51(PTPIP51)14,16,20.为了可视化 VAPB 和 PTPIP51 可能相互作用的 ERMCS 的 3D 结构,我们使用了高压冷冻,然后是冷冻取代和聚焦离子束扫描电子显微镜 (FIB-SEM)21.这种方法将接触位点保持在接近原始状态8,避免可能破坏其脆弱组织的化学固定伪影22,23,24 (补充案文,第3a节)。来自几个 COS7 细胞的小体积 ER 和线粒体被手动重建。我们将接触位点确定为 OMM 24 nm 内的 ER 膜区域,即 VAPB 的预测系留距离12,20(图。图1b,扩展数据图。1a-c和补充视频1;详见方法和补充文本第3b节)。ERMCS在FIB-SEM体积中含量很高,通常出现在线粒体嵴基部附近(图1)。1c 和扩展数据图。1c)或OMM中的收缩区域(扩展数据图。这些位点的位置与ERMCSs在线粒体能量产生和线粒体裂变中的已知作用一致25、26、27 .
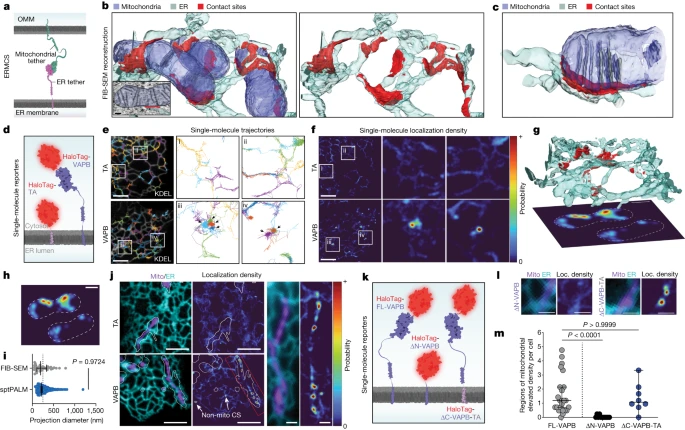
图1:内质网-线粒体接触位点的内质网系绳运动改变。
ERMCS是细胞代谢的中心枢纽,调节内质网和线粒体之间的脂质和代谢物交换。在这里,我们直接观察了活细胞中构成这些关键结构的系留机制的行为和组织。现有的细胞器间接触位点模型通常描绘了稳定地拴在一起数秒的蛋白质复合物。事实上,先前研究内质网-质膜接触位点分子活力的工作已经报告了数十秒到几分钟的停留时间50,表明整体的伪稳定性。我们使用 VAPB 系绳的高速分子跟踪和结构体积 FIB-SEM 的结果描绘了 ERMCS 的不同图景。ERMCS表现出优异的可塑性,VAPB的停留时间大约是ER-质膜接触位点蛋白质的瞬时停留时间的十倍。尽管有这种动态性,ERMCS仍将空间组织的子域保持在稳定状态,从而能够对接触点进行重塑以应对饥饿。为了强调这种亚稳态的重要性,我们发现VAPB中的致病突变破坏了这种结构和可塑性。我们在这里报告的前所未有的组分动态交换和精心维护的纳米级结构为理解ERMCS提供了一种新的范式,并将有助于指导未来旨在表征接触位点的生物学和功能的工作。
这些发现的一个关键含义是,系绳之间的相互作用可能包括接触位点整个结构中的许多快速结合和解结合事件,这与已知的VAPB对含FFAT的结合伴侣(如PTPIP51)的低亲和力一致。14,17,51).这导致整个接触位点的VAPB丰度逐渐增加,中心有一个明显的峰,由于潜在栓系分子的密度增加,再结合事件的可能性会增加。此外,预计接触位点中心的这种更高丰度的系绳将在两个膜之间提供更大的粘合力,我们可以直接观察到这种现象,即 FIB-SEM 数据集中 ERMCS 中心的负曲率区域。接触点的周围边缘区域可以作为其他ERMCS相关活动的重要中转点。支持这一观点的是,ERMCS的位置与线粒体嵴的位置和线粒体膜中的收缩位点直接相关,这些生物现象除了系绳外还需要募集特定的细胞机制25,26,27,52,53.
我们的研究结果进一步揭示了ERMCS界面表现出可塑性,能够根据不同条件调整其结构和组织。例如,可以通过像PTPIP51这样的网络共享合作伙伴的可用性来调节接触站点的大小和形状。此外,它们在营养剥夺期间变大并表现出降低的VAPB迁移率,这可能允许更有效的细胞器间代谢物转移,正如先前在经历急性饥饿的细胞的生化研究中报道的那样34,35,36,37.这些接触位点转化可能在生理上发生的一种方式是通过调节 VAPB-PTPIP51 相互作用或通过翻译后修饰任一组分来调节 ER 内 VAPB 分子之间的横向关联(如 ER -质膜接触所建议的,参见补充文本,第 2c 节,用于讨论)。事实上,有生化数据表明,这两种成分的翻译后修饰都受到精确调控14,17,51,54,55.我们在 ERMCS 和周围 ER 之间观察到的 VAPB 分子的动态交换表明,VAPB 的翻译后修饰不需要发生在接触位点内,因为 ERMCS 和 ER 定位的池将迅速平衡。进入空间受限的 ERMCS 环境可能会给与 VAPB 调节有关的大量磷酸酶和激酶带来空间挑战。我们怀疑这种策略可能是一种通用的监管机制,适用于其他系绳和接触站点。
我们对 P56S VAPB 的高速跟踪强调了 ERMCS 动态交换 VAPB 的重要性。尽管大多数P56S VAPB分子聚集或经历游离ER扩散,但一小部分但很重要的部分仍在ERMCS中。值得注意的是,这些与ERMCS相关的P56S VAPB分子破坏了接触位点的正常VAPB扩散,许多分子被困在ERMCS的多个小子域中。我们推测,这些被捕获的分子未能离开ERMCS可能导致接触位点进行正常动态重组的能力受损,无论是通过改变PTPIP51相互作用还是通过接触位点内VAPB的二聚化/横向聚集的变化(见补充文本,第2e节,供讨论)。在各种情况下,ER 和线粒体之间更稳定的界面已被证明由于 ER 到线粒体的信号传导导致线粒体功能升高11、34、54、56.因此,ERMCS中即使是一小部分P56S也可能直接导致ERMCS信号传导受损和VAPB相关ALS疾病中已知发生的氧化应激40,54.
总之,我们的结果表明,ERMCS具有不同的子域和深刻的动态性质,系绳在毫秒级的时间尺度上扩散到和扩散到站点之外,并且接触站点的大小和配置会随着不同的刺激而变化。将FIB-SEM结构数据与单颗粒跟踪相结合的未来工作有望进一步深入了解ERMCS生物学,并在分子水平上影响细胞器间通信中涉及的其他动态界面的理解。
膜接触位点
膜接触位点通常被定义为两个细胞器膜之间的紧密贴合区域。迄今为止,已经有同型(在相同的细胞器之间)和异型(在两个不同的细胞器或两种不同的膜类型之间)接触位点的例子。最初已充分研究的异型接触都涉及内质网。例如,内质网-线粒体、内质网-PM、内质网-高尔基体、内质网-过氧化物酶体和内质网脂滴 (LD) 接触。最近,发现了不涉及内质网的接触,例如:LDs-过氧化物酶体、线粒体-液泡/内体/溶酶体、线粒体-PM、线粒体-LD、线粒体-过氧化物酶体和线粒体内膜和外膜(有关迄今为止所有特征性异型接触位点的综述,请参阅18).在植物中,叶绿体与大多数其他细胞器接触20.在两个过氧化物酶体之间已经描述了非融合中间体的同型接触位点23–25或两个 LD26潜在的其他多拷贝细胞器也可能形成它们。在我们看来,作为融合中间体的细胞器之间的同型相互作用具有不同的特征,并且在操作上并不代表与这里讨论的接触位点相似,因此本综述将不涉及。还应该注意的是,无膜细胞器可以与膜状细胞器形成接触:例如,包涵体可以与LD相互作用27.然而,由于这些接触不发生在两个被膜结合的细胞器之间,它们在生理上可能非常不同,因此我们也不会在这里讨论它们。
Coming together to define membrane contact sites | Nature Communications
在真核生物中,细胞内膜界定了具有不同生化功能的细胞器。虽然几十年来,细胞器领域一直受到旨在确定每个隔室独特特征的研究的支配,但在过去的几年里,随着人们更加关注细胞器之间的相互作用及其在维持细胞稳态中的作用,该领域发生了一场革命。
1950年代后期出现了两种不同细胞器之间相互作用的已发表例子。然而,由于缺乏感知到的生理作用,因此很难将其视为一种普遍的和功能相关的现象。当时的强烈观念是,胞质溶胶的物理组织仅通过锚定和运动在细胞骨架元件上进行。此外,人们认为,两个细胞器之间疏水小分子的转移是由自由扩散的胞质蛋白催化的,并且可溶性代谢物和第二信使可以长距离传播3.这两种观点共同推迟了对两个细胞器之间膜系系重要性的理解。
当功能结合的例子变得明显时,该领域开始扩大并获得动力,这些区域被称为膜接触位点。例如,当内质网 (ER) 和线粒体之间的并列被确定为磷脂生物合成和转移的位点,几年后成为高效钙的站点2+转移5.然后将类似的作用归因于内质网-质膜 (PM) 接触位点以及ER-高尔基体接触者8.核液泡连接(NVJ)的发现及其在细胞核的零碎微自噬中的作用,后来在自噬调控中分配ER-PM接触的角色11表明接触者的作用可以是多种多样的。近年来,人们发现了更多的作用,例如ER在调节线粒体方面的作用和内体这些细胞器之间接触位点的裂变。在控制遗传中的作用,例如过氧化物酶体也被描述过。因此,细胞器是高度相互联系的,并且在接触位点的这些物理关联具有多种重要功能。
近年来,正在描述和研究其他接触者。更重要的是,以前的功能观察现在正被越来越清晰的分子理解所耦合,因为系绳,使和维持两个膜紧密接近的分子,正在被识别出来。此外,未发现的分子决定因素正与各种细胞和病理生理过程有关,证明了接触部位在正常发育和生理学中的重要性。
总之,一个全新的领域已经出现,致力于研究分子机制、细胞生物学、接触位点的生理和病理学意义。虽然现在很明显,这些排列是任何真核细胞的结构和功能的核心,并且它们正在成为细胞生物学研究的中心舞台,但同样清楚的是,就像任何新生的科学领域一样,定义和测量过程的术语和实验方法定义模糊,导致潜在的争议并阻碍知识的发展。为了克服这个问题,我们决定为该领域提供一本词典和一套实验指南。
细胞器膜接触点的功能
有人建议,如果细胞器并列具有以下特征,则可以将其定义为膜接触位点。
网络共享
我们将接触位点定义为两个双层或单层(如LD)膜结合细胞器之间的拴系接近。许多手稿定义了两个细胞器之间距离的“上限”,仍然可以定义为接触位点(通常在 10-80 nm 距离范围内,许多集中在 30 nm 范围内)。显然,间隙越窄,就越容易看到接触。然而,目前尚不清楚具体距离值是如何定义的,因为一些接触位点的容量要大得多(例如,Num1 可以跨越 300 nm28).因此,我们认为距离不能是唯一的衡量标准,并且无论距离如何,细胞器的简单并列定位都不足以被视为接触部位。在迄今(2019)为止报告的所有研究中,真正定义接触位点的是蛋白质-蛋白质或蛋白质-脂质相互作用产生的拴系力的存在。
缺乏融合
接触不包括活跃的、SNARE介导的或独立的融合过程的中间体。聚变中间体过去被称为“对接”事件,如果保留这种命名法,可以帮助保持这两个过程的区别。然而,可能存在有限的囊泡运输,但将遵循既定的机制和术语。
具体功能
解除点必须履行特定功能。由于大多数接触者最初是用内质网描述的,这突出了接触位点在脂质和 Ca 中的作用2+转移29多年来,这主导了该领域的观点。然而,现在越来越明显的是,几乎所有的细胞器都可以相互接触,而且这些接触的功能多样性要广泛得多。迄今为止,已经提出了三种类型的功能:(i)分子的特定双向传输,例如各种离子,Ca2+、脂质、氨基酸和金属。(ii) 对重塑活动很重要的信号信息或力的传递,包括细胞器生物发生、动力学、遗传、定位、裂变和自噬的调节。(iii) 反式酶(如磷脂酰肌醇 (PI) 磷酸磷酸酶 Sac1;和蛋白酪氨酸磷酸酶 1B,PTP1B)在反式中的定位),以便规范他们的活动。由于所有触点都必须具有功能,因此需要调节它们,因此触点失调会影响细胞功能,因此应通过进化压力选择触点。
https://wap.sciencenet.cn/blog-41174-1420826.html
上一篇:吸入氢气对小鼠后肢缺血再灌注损伤的保护作用
下一篇:叹为观止:AI破译出2000年前烧焦纸莎草纸文字