
KRAS基因在肿瘤发生发展过程中扮演着极为关键的角色,其突变广泛存在于多种肿瘤类型中,是重要的肿瘤驱动因素。KRAS G12C突变作为 KRAS突变的一种重要亚型,约占所有 KRAS 突变的40%[1]。Sotorasib 正是在此基础上诞生的一种 KRAS G12C 抑制剂,自问世以来,在科研领域引发了广泛且深入的研究。AbMole为全球科研客户提供高纯度、高生物活性的抑制剂、细胞因子、人源单抗、天然产物、荧光染料、多肽、靶点蛋白、化合物库、抗生素等科研试剂,全球大量文献专利引用。
一、Sotorasib(AMG-510)的作用机制
KRAS蛋白是细胞内多条重要信号通路的上游关键节点,其中 RAS-RAF-MEK-ERK(MAPK)信号通路是其发挥促癌作用的核心通路之一 。正常情况下,当细胞外生长因子与受体酪氨酸激酶结合后,通过一系列衔接蛋白的作用,促使 KRAS 蛋白结合的 GDP 转换为 GTP,KRAS 被激活进而招募并激活下游的 RAF 激酶,随后依次激活 MEK、ERK 等激酶,最终调节细胞的增殖、存活、迁移等生物学过程。在存在 KRAS G12C 突变的肿瘤细胞中,突变的 KRAS G12C 持续处于高活性状态,不断激活下游的 MAPK 信号通路,驱动肿瘤细胞的异常增殖与存活。Sotorasib(AMG-510,AbMole,M9356)能够特异性且不可逆地与 KRAS G12C 蛋白相结合,其关键在于与 KRAS G12C 突变位点的半胱氨酸残基形成共价键,这种共价结合方式具有高度的特异性,使得Sotorasib能够精准地作用于 KRAS G12C 突变体。在形成共价键后,Sotorasib 将 KRAS G12C 锁定于 GDP 结合状态,从而阻止了 KRAS 蛋白从失活的 GDP 结合形式向激活的 GTP 结合形式转换。从空间结构角度来看,Sotorasib 的结合还改变了 KRAS G12C 蛋白的构象,影响了其与下游效应分子的相互作用界面。
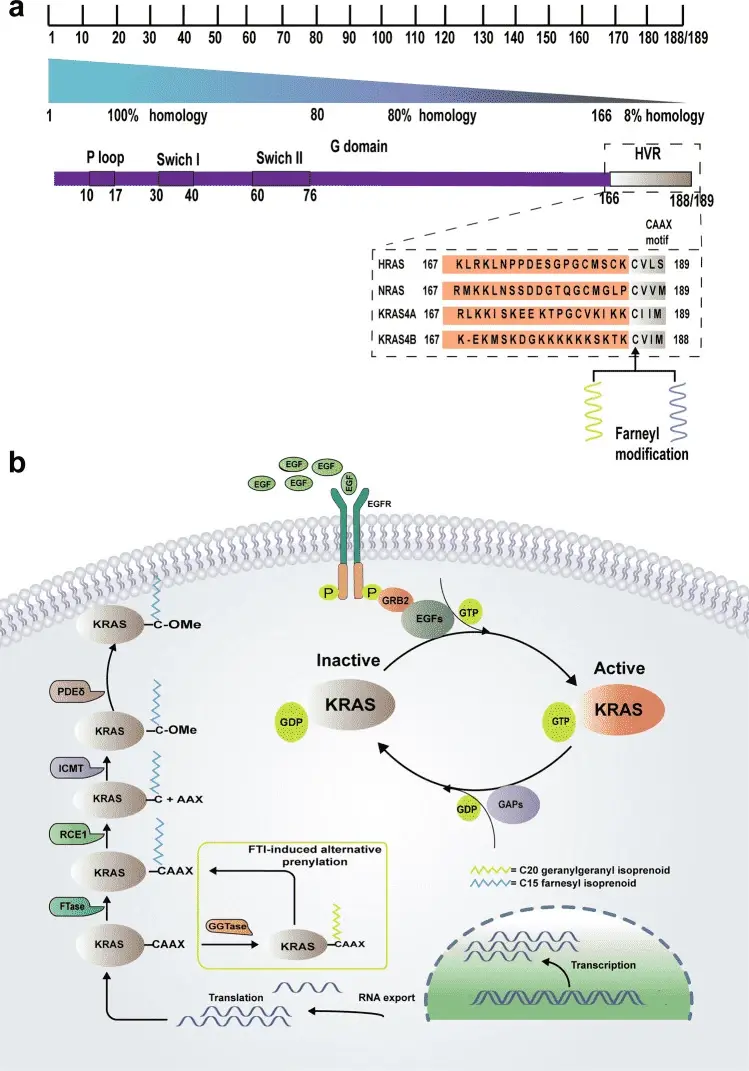
图 1. The structure and function of KRAS[2]
二、Sotorasib(AMG-510)的科研应用
1. Sotorasib(AMG-510)在肿瘤细胞系中的研究
在多种KRAS G12C突变的肿瘤细胞系研究中,Sotorasib(AMG-510,AbMole,M9356)展现出显著的生长抑制效应。以非小细胞肺癌(NSCLC)细胞系NCI-H358为例,该细胞系携带KRAS G12C突变。当使用不同浓度的Sotorasib处理 NCI-H358细胞后,通过CCK-8 细胞活力检测实验发现,细胞活力随着Sotorasib浓度的增加而逐渐降低,呈现明显的剂量依赖性。进一步通过细胞周期分析实验发现,Sotorasib能够使细胞周期阻滞在G1期,减少处于S 期和G2/M期的细胞比例,表明其抑制了细胞的增殖进程。同时,在细胞凋亡检测实验中,使用 Annexin V-FITC/PI 双染法结合流式细胞术分析发现,Sotorasib处理后的NCI-H358细胞早期凋亡和晚期凋亡细胞比例均显著增加,说明Sotorasib能够诱导肿瘤细胞发生凋亡[3]。除了NSCLC细胞系,在胰腺癌等其他携带KRAS G12C突变的细胞系如MIA PaCa-2中,也观察到类似的现象,Sotorasib能够显著降低细胞活力,诱导细胞周期阻滞和凋亡[4]。这些在肿瘤细胞系中的研究结果为Sotorasib在体内的抗肿瘤活性提供了重要的细胞学基础。2014年,AbMole的两款抑制剂分别被西班牙国家心血管研究中心和美国哥伦比亚大学用于动物体内实验,相关科研成果发表于顶刊 Nature 和 Nature Medicine。
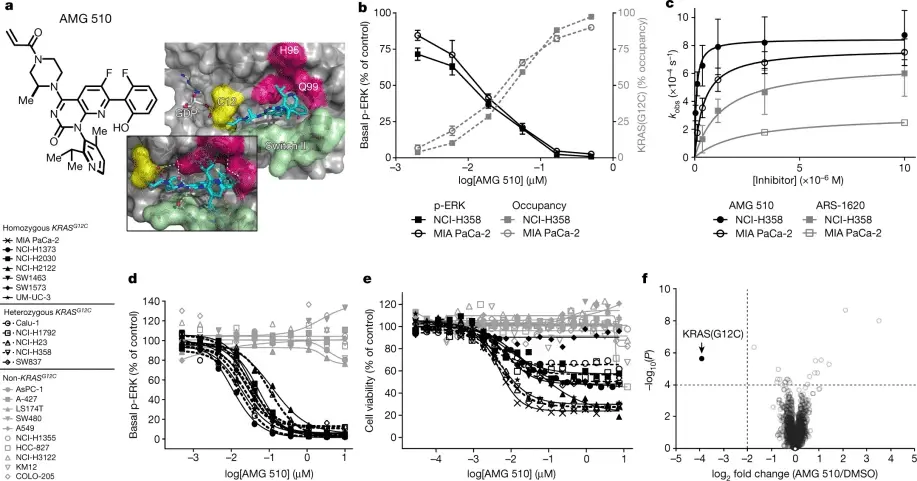
图 2. AMG 510 exploits a cryptic groove in KRAS(G12C) to enhance potency and selectivity[4]
2. Sotorasib(AMG-510)在动物模型中的研究
在KRAS G12C突变肿瘤的动物研究方面,构建小鼠模型是常用的研究手段。例如,将 KRAS G12C突变的肿瘤细胞(如NCI-H358细胞)皮下接种于免疫缺陷小鼠体内,待肿瘤生长至一定体积后,给予小鼠口服Sotorasib(30 mg/kg)[5]。结果显示,与对照组相比,Sotorasib处理组小鼠的肿瘤生长明显受到抑制,肿瘤体积增长速度显著减缓。通过对肿瘤组织进行免疫组化分析发现,Sotorasib处理组的肿瘤组织中增殖标志物Ki-67的阳性表达率显著降低,而凋亡标志物Cleaved Caspase-3的阳性表达率明显升高,进一步验证了其在体内对肿瘤细胞增殖的抑制和诱导凋亡作用。此外,在原位肿瘤模型中,如将KRAS G12C突变的肺癌细胞原位接种于小鼠肺部构建肺癌原位模型,给予Sotorasib治疗同样能够观察到肿瘤生长抑制以及肺部转移灶数量减少等现象。这些动物模型研究结果有力地证明了Sotorasib(AMG-510,AbMole,M9356)在体内具有良好的抗肿瘤活性[6]。
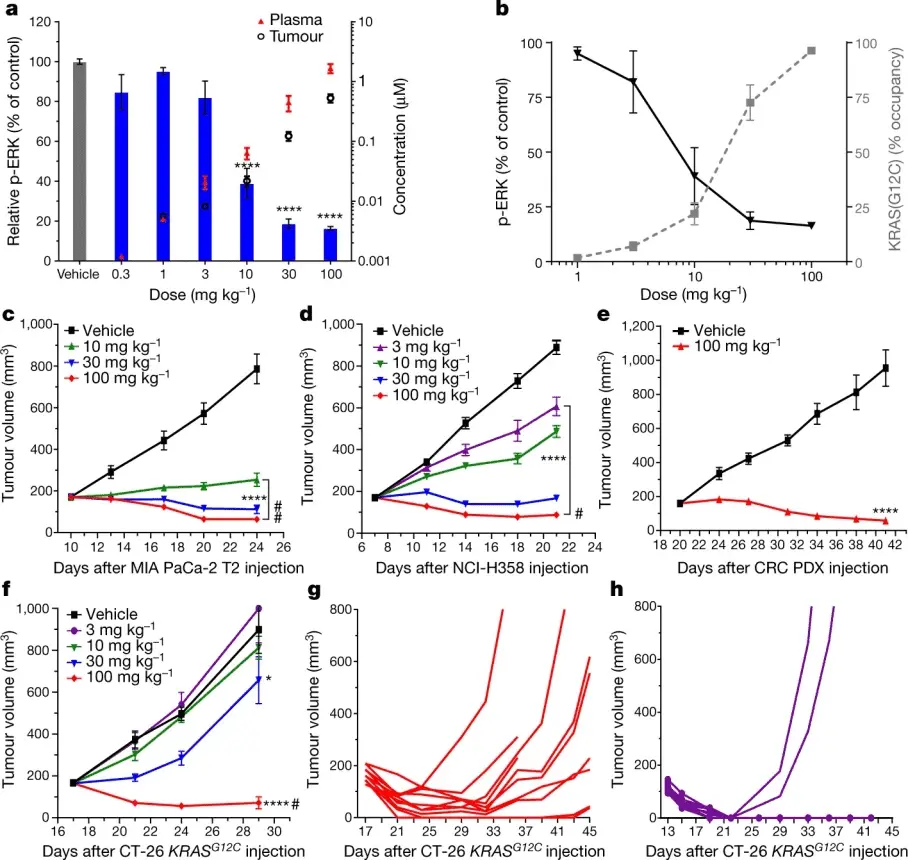
图 3. AMG 510 inhibits ERK phosphorylation and growth of KRASG12C-mutant tumours in vivo[4]
3. Sotorasib(AMG-510)与人源化单抗和其他抑制剂的联合使用研究
Sotorasib(AMG-510,AbMole,M9356)可以与多种抑制剂或人源化单抗联用,例如MET信号通路抑制剂--Crizotinib [7]。有研究表明在KRAS G12C 被抑制后,可能会通过反馈激活MEK - ERK信号通路。因此,同时使用Sotorasib和Crizotinib能够阻断这一潜在的逃逸通路[7]。在结直肠癌(CRC)的研究中发现,EGFR 通路的上调是导致对Sotorasib等KRAS G12C抑制剂产生耐受性的关键机制之一。人源化单抗中的Panitumumab作为EGFR抑制剂,为 Sotorasib的组合方案提供了一种极具潜力的方向[8]。此外,Sotorasib还可与免疫检查点抑制剂如Atezolizumab(AbMole,M6101)、BMS-1(AbMole,M8959)联合使用。通过激活免疫和肿瘤生长抑制之间的协同,有效抑制剂多种肿瘤细胞的生长[9]。
三、范例详解
iScience. 2023 Jan 28;26(2):106080.
蛋白质组学国家重点实验室、中国科学院植物研究所的实验人员在上述文章中探究了KRAS G12C 共价抑制剂AMG510的蛋白修饰谱,并通过开发两种分析方法(直接Profiling和基于泛 AMG510抗体的肽段免疫沉淀),系统性鉴定了AMG510的靶向及脱靶修饰位点,揭示其潜在的生物学效应和毒理学意义。上述论文的研究核心—KRAS G12C 共价抑制剂AMG510(Sotorasib,AbMole,M9356)由AbMole提供。此外,实验人员还使用了AbMole的MRTX849(Adagrasib,AbMole,M9428),另一种与AMG510结构相似的 KRAS G12C共价抑制剂,以验证泛 AMG510 抗体的靶向性,即该抗体仅识别 AMG510 修饰的肽段,不识别 MRTX849 修饰或未修饰的肽段[10]。
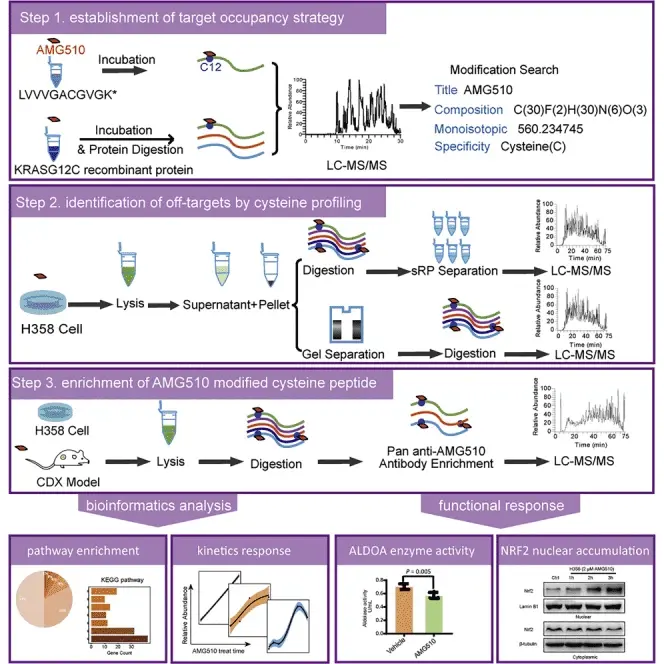
图 4. Global profiling of AMG510 modified proteins identified tumor suppressor KEAP1 as an off-target[10]
AbMole是ChemBridge中国区官方指定合作伙伴。
参考文献及鸣谢
[1] J. Moldvay, J. Tímár, KRASG12C mutant lung adenocarcinoma: unique biology, novel therapies and new challenges, Pathology oncology research : POR 29 (2023) 1611580.
[2] L. Huang, Z. Guo, F. Wang, et al., KRAS mutation: from undruggable to druggable in cancer, Signal transduction and targeted therapy 6(1) (2021) 386.
[3] Karen Rex, Anne Y. Saiki, Tyler Holt, et al., Abstract 1057: Combination of the KRASG12C inhibitor sotorasib with targeted agents improves anti-tumor efficacy in KRAS p.G12C cancer models, Cancer research 81(13_Supplement) (2021) 1057-1057.
[4] J. Canon, K. Rex, A. Y. Saiki, et al., The clinical KRAS(G12C) inhibitor AMG 510 drives anti-tumour immunity, Nature 575(7781) (2019) 217-223.
[5] M. B. Ryan, F. Fece de la Cruz, S. Phat, et al., Vertical Pathway Inhibition Overcomes Adaptive Feedback Resistance to KRAS(G12C) Inhibition, Clinical cancer research : an official journal of the American Association for Cancer Research 26(7) (2020) 1633-1643.
[6] D. J. Sisler, T. K. Hinz, A. T. Le, et al., Evaluation of KRAS(G12C) inhibitor responses in novel murine KRAS(G12C) lung cancer cell line models, Frontiers in oncology 13 (2023) 1094123.
[7] Shinichiro Suzuki, Kimio Yonesaka, Takeshi Teramura, et al., KRAS Inhibitor Resistance in MET-Amplified KRASG12C Non–Small Cell Lung Cancer Induced By RAS- and Non–RAS-Mediated Cell Signaling Mechanisms, Clinical Cancer Research 27(20) (2021) 5697-5707.
[8] Y. Kuboki, M. Fakih, J. Strickler, et al., Sotorasib with panitumumab in chemotherapy-refractory KRAS(G12C)-mutated colorectal cancer: a phase 1b trial, Nature medicine 30(1) (2024) 265-270.
[9] P. Du, L. Liu, T. Hu, et al., Integrative Analysis of Pharmacokinetic and Metabolomic Profiles for Predicting Metabolic Phenotype and Drug Exposure Caused by Sotorasib in Rats, Frontiers in oncology 12 (2022) 778035.
[10] Y. Wang, B. Zhong, C. Xu, et al., Global profiling of AMG510 modified proteins identified tumor suppressor KEAP1 as an off-target, iScience 26(2) (2023) 106080.
转载本文请联系原作者获取授权,同时请注明本文来自王开来科学网博客。
链接地址:https://wap.sciencenet.cn/blog-3646309-1507277.html?mobile=1
收藏