
撰文 | 李欣茜 张婷 郑宇含 张彦康 李雨
编辑 | 孟美瑶
校对 | 张婷

背景介绍
健康寿命是指人们保持良好健康状态的生命阶段,改善健康寿命对于改善老年人群生活质量、延长预期寿命至关重要。目前,65岁及以上的成年人中有35%到40%被归类为肥胖人群。肥胖和衰老均会增加慢性疾病的风险,因此深入理解二者如何协同影响健康寿命与预期寿命,对开发有效治疗策略具有关键意义。
成纤维细胞生长因子21(FGF21)是FGF家族的非典型成员,是一种内分泌激素,主要由肝细胞分泌进入循环系统以响应多种细胞应激。除肝脏外,FGF21还可在胸腺上皮细胞、脂肪细胞、肌肉和胰腺局部合成,这些局部产生的FGF21可能发挥自分泌和旁分泌作用。FGF21通过由FGF受体1c(FGFR1c)与辅助受体β-Klotho组成的异源二聚体复合物进行信号传导。虽然FGFR1c在多种组织中广泛表达,但β-Klotho主要在大脑、肝脏和脂肪组织中表达,因此这些部位是FGF21的主要作用靶点。在小鼠中的研究表明,在中枢神经系统(CNS)和脂肪组织中,FGF21对于增强能量消耗和改善胰岛素敏感性至关重要。目前的研究表明FGF21能显著改善小鼠和人类代谢性疾病(如2型糖尿病和非酒精性脂肪肝病),其已成为备受关注的潜在治疗药物,但FGF21作用于多器官的具体分子机制仍未阐明。
目前的研究普遍认为FGF21在衰老进程中具有积极作用,并将FGF21定义为“促长寿激素”,然而很少有研究直接探讨FGF21对寿命的影响。此前有研究表明,肝脏持续表达FGF21能显著延长小鼠寿命,但该效应被归因于能量消耗增加以及对生长通路(包括胰岛素样生长因子(IGF)和生长激素(GH)信号传导)的调节。然而,在这些研究从小鼠发育期开始便持续性表达FGF21,会导致小鼠发育异常和侏儒症等副作用。因此,在正常小鼠中、以及更接近人类疾病和衰老状态的条件下重新评估FGF21的作用尤为重要。此外,有研究表明FGF21在调节适应性饥饿反应中发挥关键作用,并且这一过程与寿命延长相关。据报道,FGF21缺失会逆转蛋白质限制饮食带来的寿命延长效应。由于全球大多数人口的代谢衰老与营养过剩有关,因此进一步研究FGF21在衰老与肥胖状态下的作用机制具有重要意义。
在本篇文章中,研究人员构建了脂肪细胞特异性过表达FGF21的诱导型转基因(TG)小鼠模型,探究在高脂饮食(HFD)造成的代谢应激条件下,局部过表达FGF21是否能够影响小鼠全身代谢状态和寿命。研究结果表明,FGF21水平的升高能够维持脂肪组织健康状态和降低神经酰胺水平,进而延长健康寿命和延长寿命。此外,本文揭示内脏脂肪组织是介导FGF21全身代谢益处的关键靶点之一,这为衰老肥胖背景下的代谢缺陷及炎症治疗提供了新的策略。

敲黑板啦!
1.过表达FGF21可延长寿命,不依赖于生长/发育调控作用
2.过表达FGF21可预防HFD饮食下衰老小鼠的肥胖、肝脂肪变性和胰岛素抵抗
3.过表达FGF21可提高HFD饮食下年轻小鼠的能量消耗,但不影响产热水平
4.FGF21独立于脂联素的作用来降低内脏脂肪中神经酰胺含量
研究结果
1.构建脂肪细胞特异性过表达FGF21小鼠模型
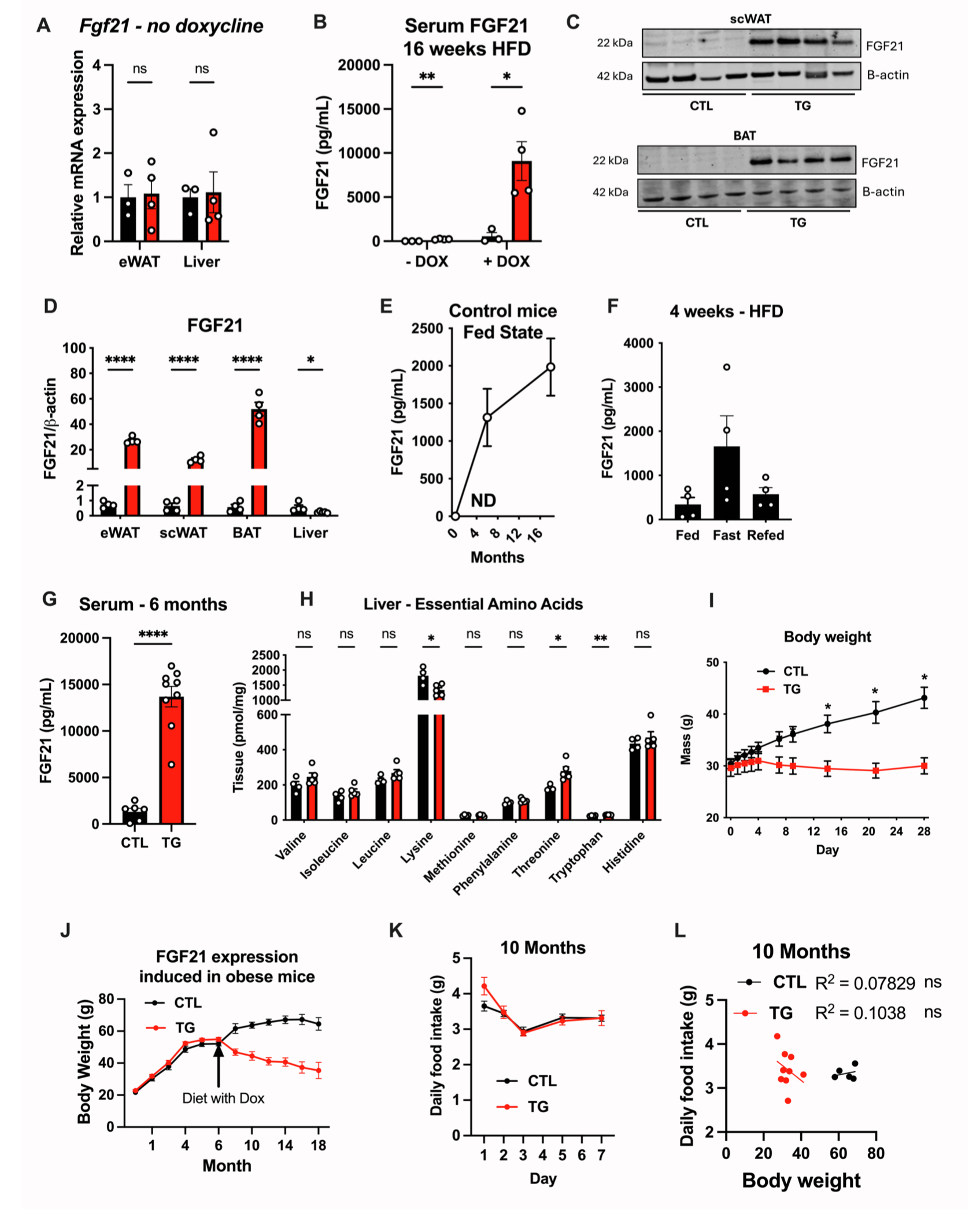
为了探究FGF21表达对衰老小鼠寿命的影响,研究人员使用Adiponectin驱动的rtTA来诱导FGF21的过表达,该小鼠在Dox(多西环素)的诱导下可在脂肪组织中特异性过表达FGF21,即TG小鼠。结果显示,在HFD-Dox饮食诱导下,FGF21特异性在eWAT、scWAT和BAT中高表达,而肝脏中FGF21的mRNA表达和蛋白表达水平下降,肝脏中调控FGF21表达和分泌的相关基因(如Xmox1、Atf4和Chop)表达无明显差异(图1A-1F,S1A-S1D)。且TG小鼠血清FGF21含量显著升高,约是禁食应激下血清FGF21的5倍(图S1E-S1G)。为了明确TG小鼠血清FGF21含量的升高是由于脂肪组织FGF21的过表达,研究人员将TG小鼠与FGF21 KO小鼠(全身性敲除FGF21小鼠)杂交,结果显示,脂肪组织特异性过表达FGF21显著提高了血清中FGF21的水平(图1C)。TG小鼠模型也避免了低蛋白饮食对肝脏中由FGF21介导的应激刺激(图S1H)。
以上结果表明,在TG小鼠模型中脂肪组织特异性过表达FGF21可提高血清中FGF21含量,且不依赖于肝脏。先前许多研究利用遗传学或药理学手段在短期内探究了FGF21的作用机制,而TG小鼠模型是第一个在整个衰老过程中过表达FGF21的小鼠模型。因此,TG小鼠的潜在表型变化都可能归因于血清中脂肪组织来源FGF21含量的增加。
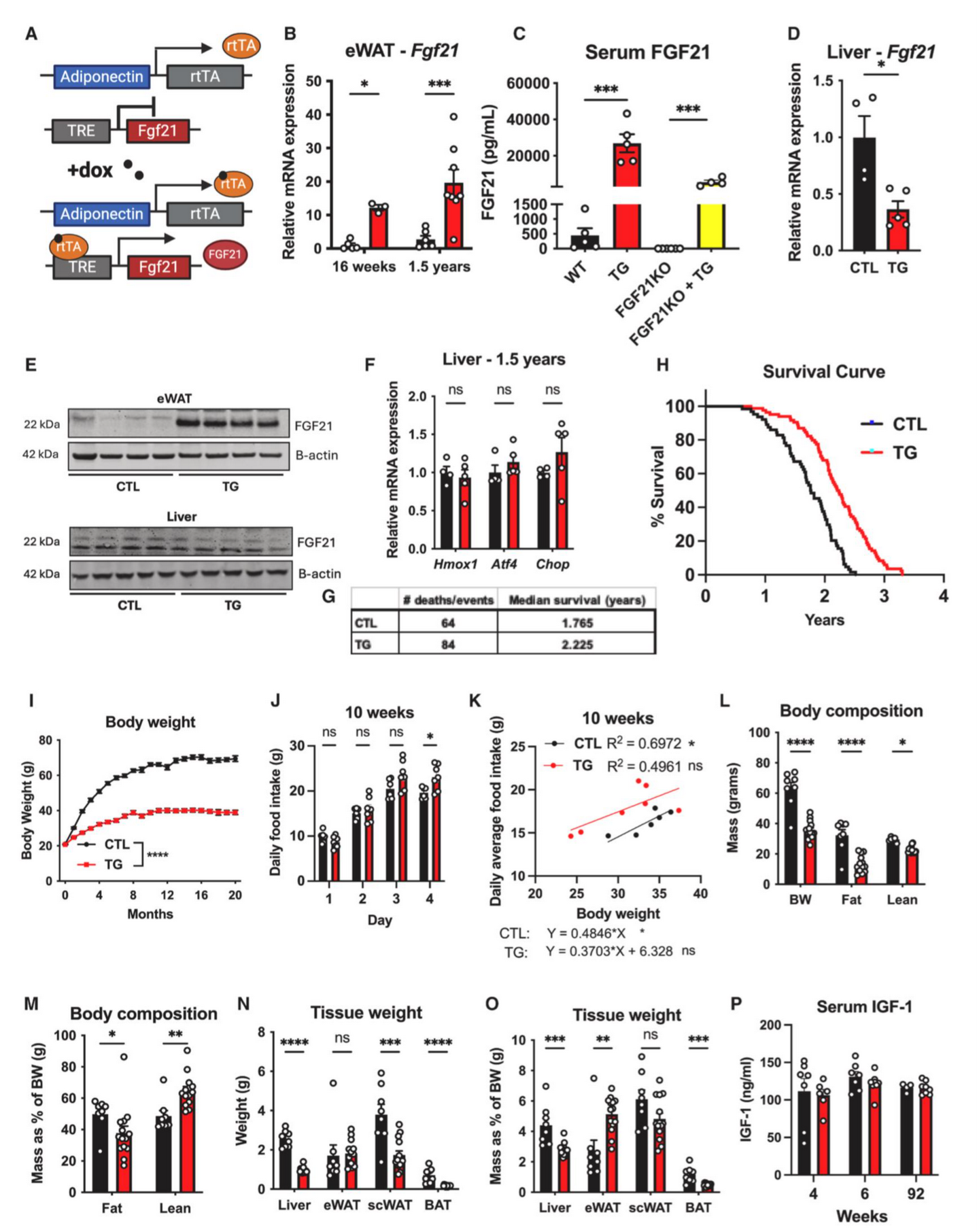 图1.血清中脂肪来源FGF21可延长寿命
图1.血清中脂肪来源FGF21可延长寿命
图S1.血清中脂肪来源FGF21可延长寿命
2.脂肪组织过表达FGF21可延长HFD小鼠寿命
接下来,研究人员给10-12周龄雄鼠饲喂HFD-Dox饮食,探究FGF21过表达是否可延长HFD喂养期间小鼠的寿命,结果显示,与对照小鼠相比,TG小鼠的寿命中位数显著延长(1.765年延长至2.225年),部分小鼠寿命长达3.30年(图1G-1H)。这些数据表明,脂肪组织特异性过表达FGF21能够有效减轻衰老的影响,从而延长寿命。
3.过表达FGF21可抵抗小鼠肥胖
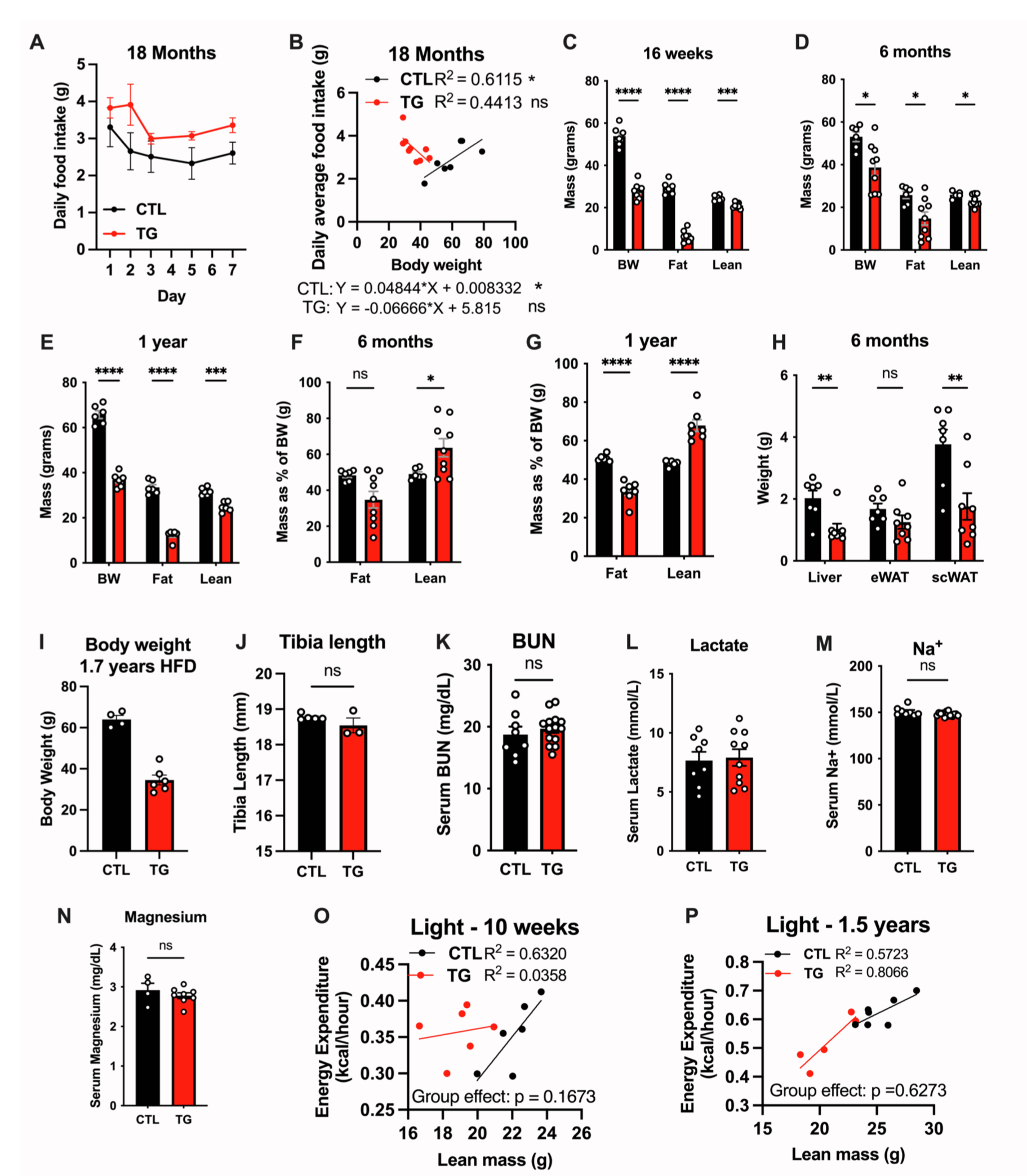
肥胖和脂肪组织功能失调会导致机体葡萄糖和脂质代谢紊乱,进而导致衰老相关疾病。结果显示,在HFD-Dox饮食,TG小鼠体重显著轻于对照小鼠,且这一现象持续到老年(图S1J)。在开始饲喂HFD-Dox饮食以及饲喂10个月和18个月时,TG小鼠的摄食量与对照小鼠相比均无显著差异,而对对照小鼠的TG小鼠的摄食量与体重之间进行回归性分析,结果显示TG小鼠在整个周期中的摄食量与体重无相关性,而对照小鼠在刚开始饲喂HFD-Dox饮食以及HFD-Dox饲喂18个月时显示出摄食量与体重的显著正相关,而在HFD-Dox饲喂10个月时无这一现象(图1J、1K、S1K、S1L、S2A和S2B)。对照小鼠的摄食量与体重增长一致,而TG小鼠在摄食量无明显变化的情况下保持较低的体重,这可能与脂肪组织FGF21的过表达相关。有研究报道FGF21是一种“饥饿激素”,促使小鼠的蛋白质摄入量增加(小编注:有研究表明,FGF21可作用于下丘脑和腹侧被盖区(VTA)的多巴胺神经元,VTA区是大脑奖赏系统的核心区域。在正常饮食下,多巴胺神经元对碳水化合物反应更强,而在低蛋白饮食下,FGF21信号促进VTA区多巴胺神经元的活性,使其对蛋白质刺激更加敏感,从而增加蛋白质摄入量。在FGF21 KO小鼠和神经元特异性敲除βKlotho(FGF21受共体)小鼠中这一现象消失。(Md Shahjalal H Khan, et al.Mol Metab. 2025 Jan:91:102068.) )。在摄入足够蛋白质的情况下,FGF21通过增加能量消耗而非热量限制来减轻体重。
体成分分析结果显示,在HFD-Dox饲喂16周和1.5年时,与对照小鼠相比,TG小鼠的Fat mass和Lean mass显著下降,而以体重归一化后,TG小鼠的Fat mass水平下降,Lean mass水平升高(图1L-1M,S2C-S2G),同时TG小鼠肝脏、scWAT和BAT重量下降,eWAT重量没有差异(图1N和S2H)。
图S2.脂肪特异性过表达FGF21并未引起生长发育的信号调控,但在年轻时增加了小鼠的能量消耗
4.FGF21的延寿作用不依赖于生长信号传导
FGF21的抗衰老作用涉及生长激素(GH)和胰岛素样生长因子-1(IGF-1)信号传导,在骨稳态调控和细胞生长方面发挥着重要作用。因此,研究人员想要探究脂肪组织FGF21是否通过IGF-1信号延长衰老。结果显示,与对照小鼠相比,TG小鼠血清IGF-1水平无明显差异(图1P)。随后研究人员进一步评估了FGF21对骨稳态调控方面的影响,结果显示,与HFD-Dox饲喂的衰老对照小鼠相比,TG小鼠的胫骨长度、骨小梁矿物质密度或皮质组织矿物质密度均无显著差异(图2A、2B、S2I和S2J),且TG小鼠血清尿素氮(BUN)、乳酸、钠和镁的水平均无显著差异(图S2K–S2N)。这些发现表明,脂肪组织过表达FGF21导致的寿命延长并非主要由生长/发育的调控引起(小编注:本文在背景中描述到,先前研究表明在肝细胞中特异性过表达FGF21可延长小鼠寿命,其机制依赖于生长/发育途径,该小鼠模型在发育期开始便持续性表达FGF21,这会导致小鼠发育异常和侏儒症等副作用。本篇文章是在小鼠成年后(10-12周)利用Dox诱导脂肪组织FGF21过表达,不影响生长/发育,这更接近于人类疾病和衰老状态)。
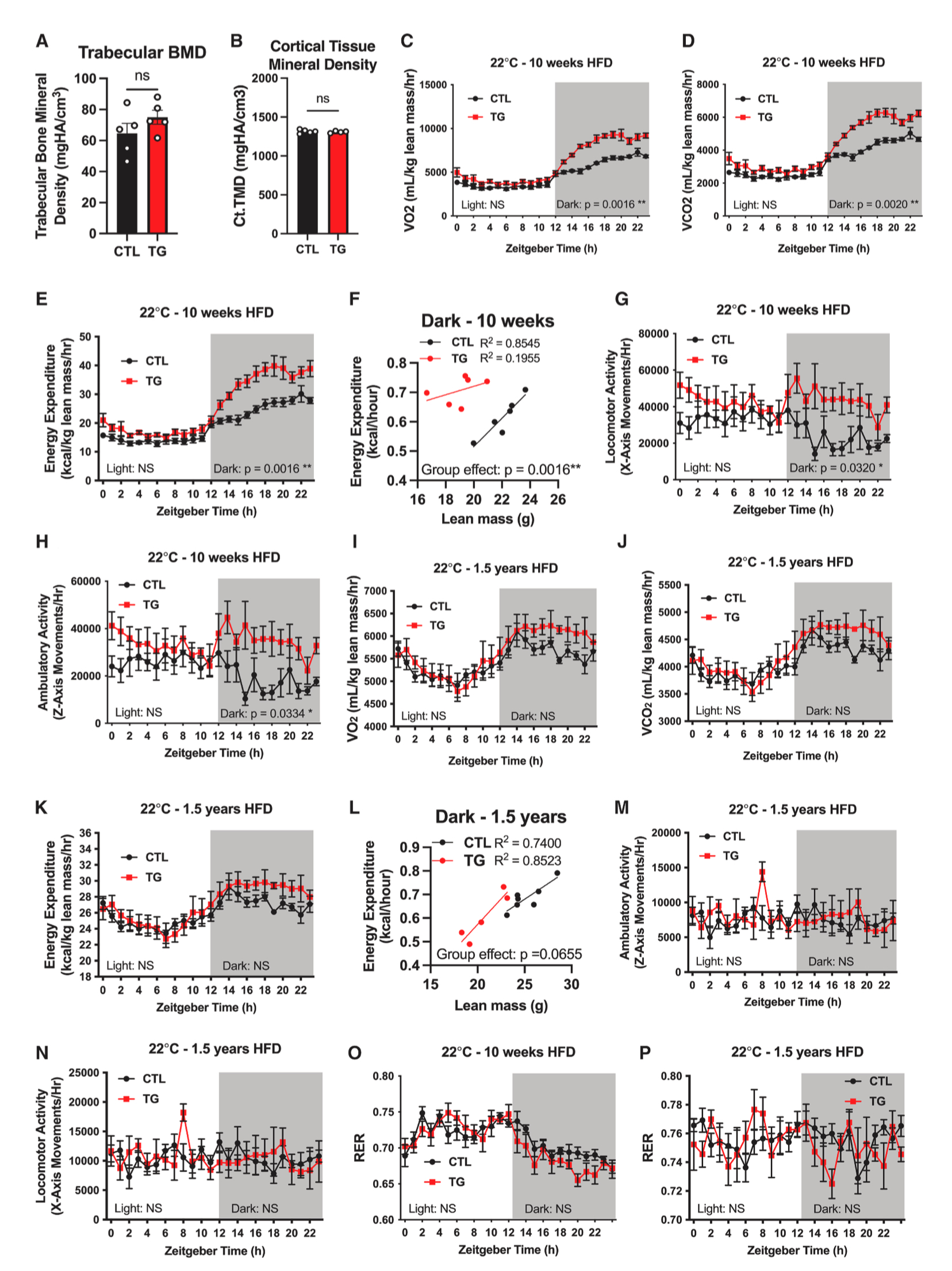
图2.脂肪特异性过表达FGF21并未引起生长发育的信号调控,但在年轻时增加了小鼠的能量消耗
5.过表达FGF21可提高年轻HFD小鼠的能量消耗
由于TG小鼠食物摄入量无明显差异,而体重减轻,因此,研究人员监测了HFD-Dox饲喂10周和1.5年的衰老对照小鼠和TG小鼠的能量消耗,结果显示,以Lean mass归一化,与HFD-Dox饲喂10周的对照小鼠相比,黑暗周期中TG小鼠耗氧量、二氧化碳产生量、能量消耗、活动量显著增加,ANCOVA分析也表明TG小鼠能量消耗显著升高,在光照周期中,TG小鼠的能量消耗无明显差异(图2C-2H,S2O)。然而,在HFD-Dox饲喂1.5年后,TG小鼠的氧气消耗量、二氧化碳产生量、能量消耗和活动量无明显差异(图2I-2N,S2P)。此外,在HFD-Dox饲喂10周或1.5年后,于对照小鼠相比,TG小鼠的RER(呼吸交换比)无明显差异,表明对照小鼠和TG小鼠利用碳水化合物或脂质功能的比例相当(图2O和2P)。总之,这些结果表明,在年轻的时候,FGF21通过促进机体能量消耗和活动量来产生显著的代谢益处,而在衰老小鼠中,FGF21介导的能量消耗可能随着年龄增长被代偿性抵消。
6.过表达FGF21可改善衰老小鼠葡萄糖代谢稳态
衰老会引起胰岛素抵抗。接下来,研究人员探究了HFD-Dox饮食下年轻和衰老TG小鼠的胰岛素敏感性和糖稳态变化。GTT和ITT结果显示,与HFD-Dox饲喂16周和1.5年的对照小鼠相比,TG小鼠的胰岛素敏感性和糖稳态均显著改善(图3A-3D),且在有/无葡萄糖刺激的条件下,TG小鼠血清胰岛素水平显著下降,eWAT和scWAT中胰岛素信号途径相关蛋白pAKT/AKT水平升高(图3E-3G)。这些结果表明脂肪组织过表达FGF21提高了小鼠胰岛素敏感性和血糖稳态。
H&E染色结果显示,与HFD-Dox饲喂1.5年的对照小鼠相比,TG小鼠eWAT和scWAT脂肪细胞面积减小,且冠状结构减少(图3I)。过氧化物酶体增殖物激活受体γ(PPARγ)是脂肪细胞中脂肪生成和胰岛素敏感性的主要调节因子。研究人员发现与HFD-Dox饲喂1.5年的对照小鼠相比,TG小鼠eWAT和scWAT中Pparg基因表达显著上调,同时eWAT中脂联素基因表达和血清中脂联素含量也显著上调,(图3J-3L),高水平的脂联素是健康脂肪组织和胰岛素敏感性的标志。总之,这些结果表明,脂肪组织过表达FGF21对eWAT和scWAT功能以及机体葡萄糖代谢稳态具有显著且持续的改善作用。
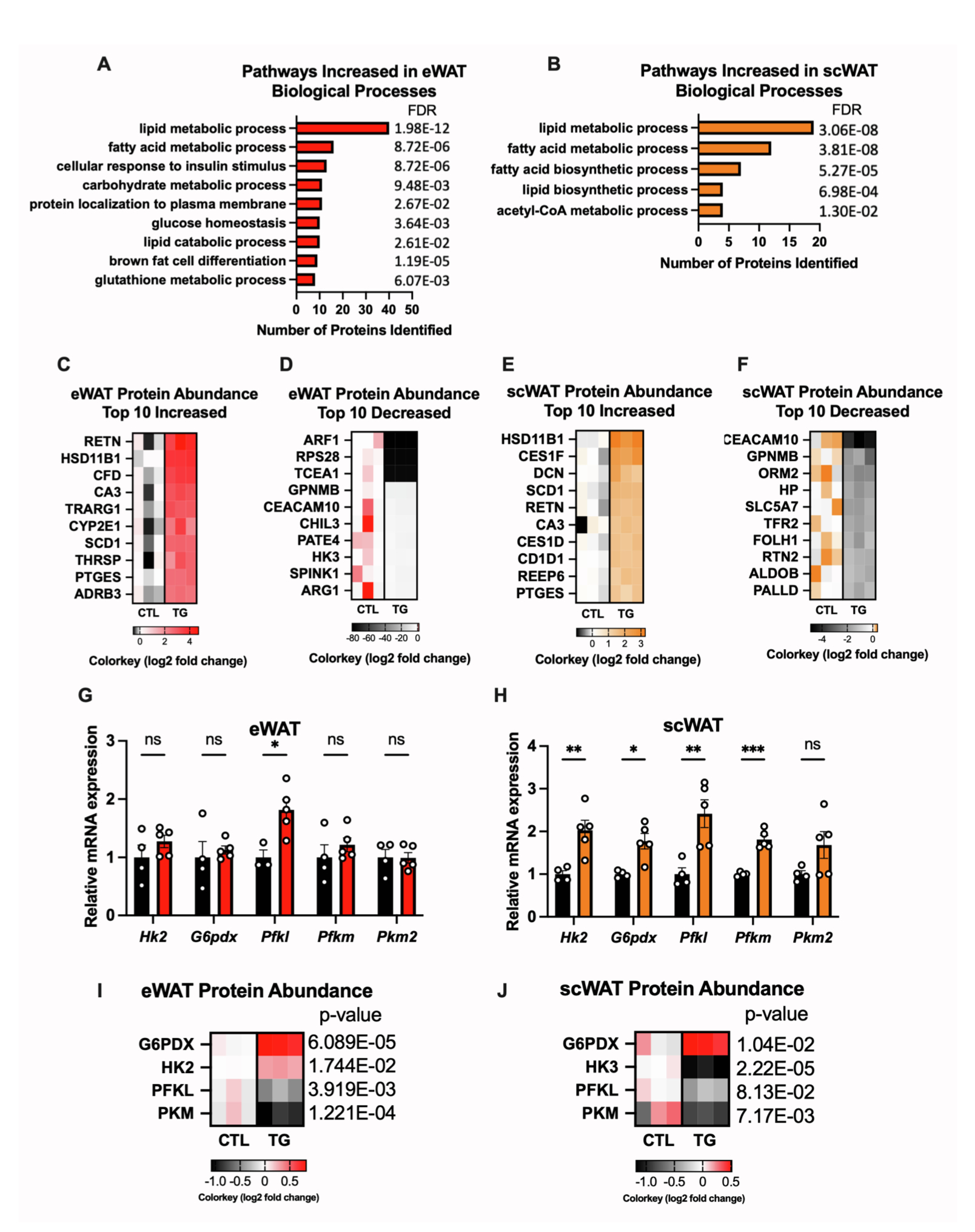
为了进一步探究FGF21影响脂肪细胞功能的分子机制,研究人员对HFD-dox 饲喂1.5年的衰老对照小鼠和TG小鼠eWAT和scWAT进行定量蛋白质组学分析和qPCR实验分析,结果显示,脂肪组织过表达FGF21对eWAT和scWAT的蛋白质组产生显著的重塑,在eWAT中变化更加显著(图3M,S3A-S3F)。其中,与对照小鼠相比,TG小鼠eWAT和scWAT中胰岛素信号途径相关蛋白和基因表达显著上调,包括关键代谢调节因子Glut4(Slc2a4)、Pck1、Fasn和Acaca(图3N- 3Q),并且TG小鼠eWAT组织中脂联素和GLUT4转运调节因子(Trarg1)表达显著上调,而在scWAT中无显著差异(图3O和3Q),此外TG小鼠scWAT中调控糖酵解途径相关基因表达显著上调,而eWAT中仅有6-磷酸果糖激酶(Pfkl)表达增加,但糖酵解限速酶如PFKL和PKM蛋白水平在TG小鼠的eWAT和scWAT中均显著下降(图S3G-S3J)。总之,这些结果表明,在肥胖-衰老诱导的代谢应激下,脂肪组织过表达FGF21对eWAT和scWAT胰岛素敏感性产生了深刻的影响。
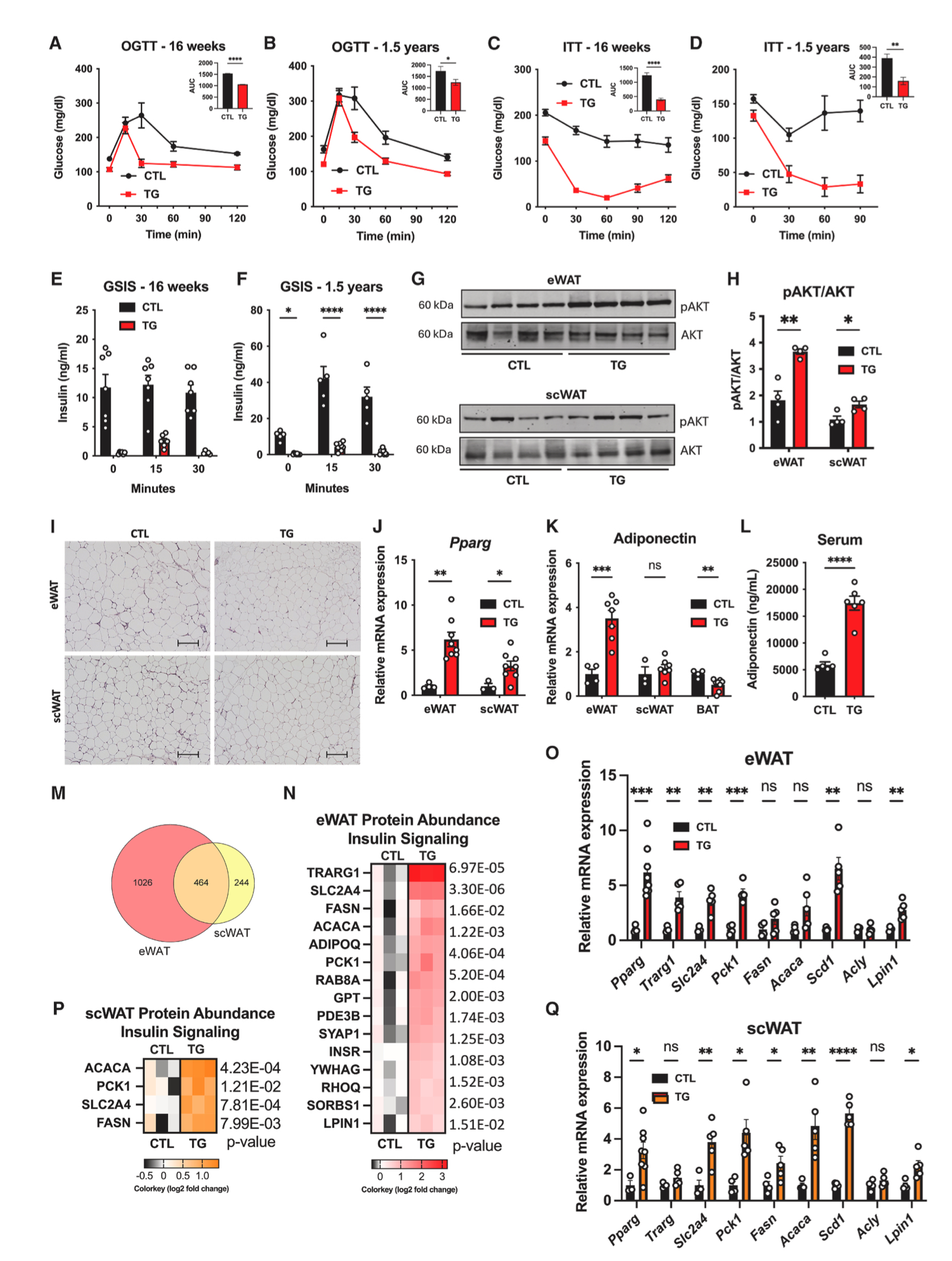
图3.脂肪组织特异性过表达FGF21可改善HFD饮食下衰老小鼠的脂肪组织功能和机体胰岛素敏感性
图S3.脂肪组织特异性过表达FGF21可改善HFD饮食下衰老小鼠的脂肪组织功能和机体胰岛素敏感性
7.过表达FGF21可促进HFD小鼠脂肪细胞的脂生成
在血糖水平升高的情况下,胰岛素可激活胰岛素信号途径上调脂肪生成相关酶活性。相反,肥胖和胰岛素抵抗会抑制脂肪细胞的脂肪生成途径。由于脂肪细胞特异性过表达FGF21小鼠的胰岛素敏感性增强,因此研究人员进一步探究了过表达FGF21是否能促进脂肪细胞的脂肪生成和脂质储存。对HFD饲喂1.5年的TG小鼠eWAT和scWAT组织蛋白质组学中表达上调的蛋白进行GO分析,结果显示FGF21过表达上调了eWAT和scWAT中脂肪生成相关蛋白,如ACACA、FASN、SCD1和ACLY(图4A和4B)。在这些蛋白中,只有Scd1的基因表达在eWAT中显著增加,Scd1、Fasn和Acaca的基因表达在scWAT中均显著增加(图3O和3Q)。此外,将eWAT重量和scWAT重量以体重归一化,结果显示TG小鼠的eWAT重量显著高于对照小鼠,而scWAT重量无显著差异(图1O)。这些结果表明过表达FGF21可促进小鼠脂肪组织的脂肪生成水平。
接下来,研究人员探究了FGF21对脂肪组织脂解的影响。首先研究人员对HFD-Dox喂养8周的对照小鼠和TG小鼠腹腔注射β3-肾上腺素受体激动剂(CL316,243)以刺激脂肪组织的脂解,结果显示TG小鼠血清中甘油和NEFAs水平显著低于对照小鼠(图4C和4D)。同时,HFD-Dox喂养1.5年的衰老TG小鼠血清中NEFA水平显著低于对照小鼠(图4E)。这些结果表明过表达FGF21可抑制小鼠脂肪组织的脂解水平。
总之,这些结果表明脂肪组织过表达FGF21可能通过促进脂肪生成途径,抑制脂解(小编注:可能过表达FGF21促进脂肪组织脂生成途径的同时,还增强脂肪代谢。本文并没有实验证明脂肪组织的脂解水平,只是通过检测血清NEFA和甘油水平表明TG小鼠脂肪组织脂解抑制,推测可能脂肪组织内部利用了脂肪酸进行能量消耗,并没有将脂肪酸释放到血清中。这提高了机体能量消耗,使TG小鼠抵抗HFD饮食诱导的肥胖,具体表现为:1. 与对照小鼠相比,在TG小鼠eWAT和scWAT在脂生成增强的同时,脂肪细胞面积更小,且eWAT重量占体重的比例升高,scWAT重量占体重的比例无差异,提示TG小鼠eWAT和scWAT组织中脂肪细胞增殖水平升高,这是脂肪组织代谢增强的一种特征;2. eWAT和scWAT中脂解抑制,避免了脂质在其他组织(如肝脏)的异位沉积,减少了机体脂毒性;3. 年轻时TG小鼠机体能量消耗增加),进而改善机体脂质稳态。
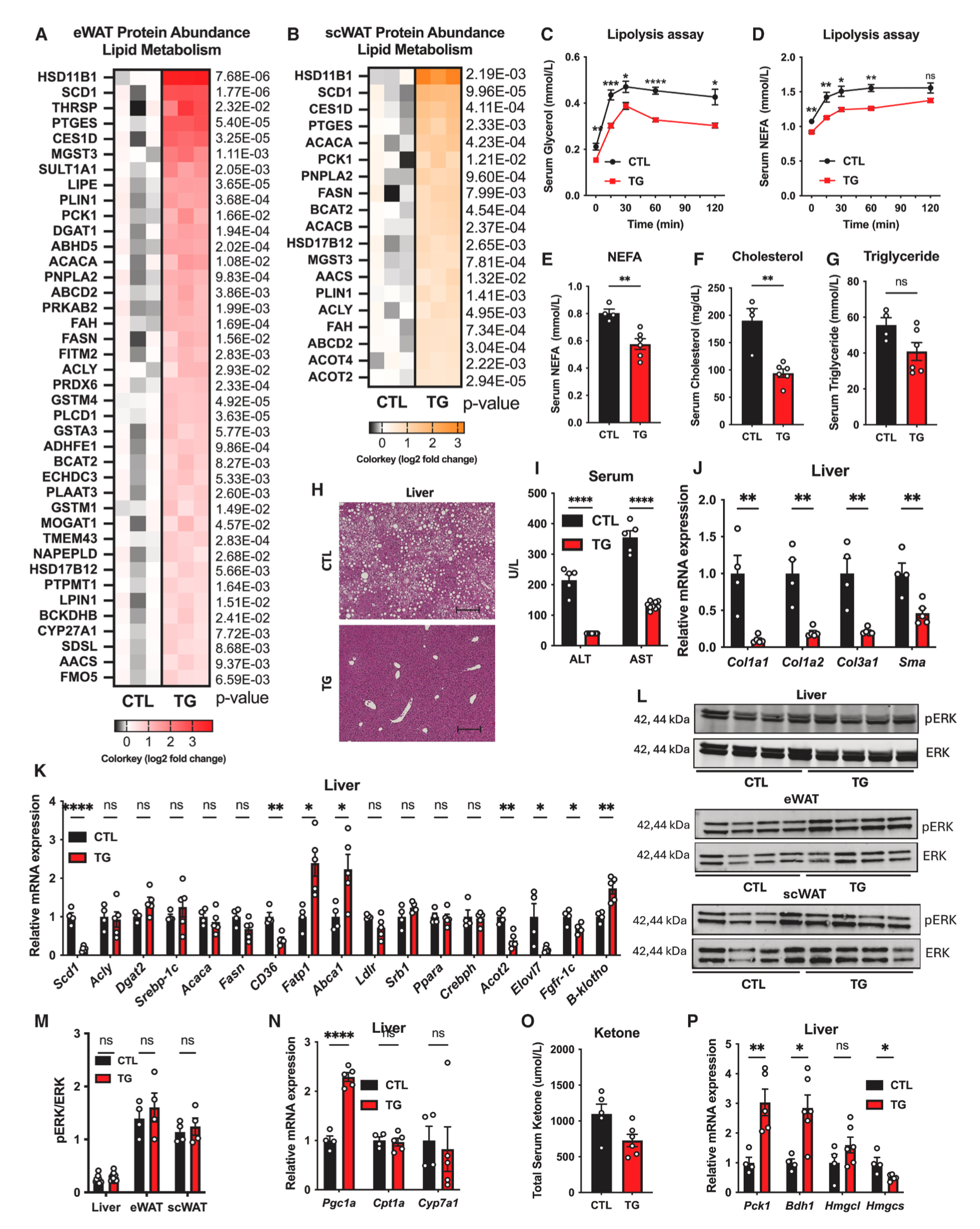
图4.脂肪细胞特异性过表达FGF21可维持白色脂肪组织的脂质代谢,并防止高脂饮食小鼠在饲养1.5年后出现肝脂肪变性
8.FGF21可改善HFD喂养的衰老小鼠肝脂肪变性现象
许多研究表明,衰老过程中,脂肪组织功能障碍可导致肝脏脂质异位沉积、肝脂毒性和代谢功能障碍相关的脂肪肝病(MAFLD)。因此研究人员想要探究脂肪组织过表达FGF21对肝脏的影响。结果显示,与HFD-Dox饲喂1.5年的衰老对照小鼠相比,衰老TG小鼠血清胆固醇和甘油三酯水平显著降低,肝脏脂质异位沉积水平下降(图4F-4H)。在HFD-Dox饲喂16周的TG小鼠中,肝脏损伤水平与对照小鼠无显著差异,而在HFD-Dox饲喂6个月和1.5年的TG小鼠中,肝脏损伤水平显著低于对照小鼠(S4A和S4B)。此外,HFD-Dox饲喂1.5年的TG小鼠肝脏中纤维化相关基因表达显著低于对照小鼠(图4J)。
先前研究表明肝脏FGF21可调控肝脏糖异生、脂肪酸氧化和酮体生成途径。接下来研究人员探究了脂肪组织过表达FGF21对肝脏中相关基因表达和信号途径的影响。结果显示,与HFD-Dox饲喂1.5年的对照小鼠相比,TG小鼠肝脏中除Scd1外其他调控脂肪生成的经典基因表达无显著差异,负责脂肪酸转运的相关基因Cd36显著下降而Fatp1表达显著上调,受FGF21调控的基因Abca1表达上调,调控FGF21表达的转录因子PPARα和Crebph表达无显著变化,而其他PPARα靶点Acot2和elov17表达显著下降(图4K)。FGF21可通过FGFR1c/β-klotho信号通路激活丝裂原活化蛋白激酶(MAPK)和细胞外信号调节激酶(ERK)通路,进而调节葡萄糖摄取和能量代谢。结果显示,与对照小鼠相比,TG小鼠肝脏中Fgfr-1c基因表达下降,β-klotho基因表达显著上调(图4K),ERK信号途径在TG小鼠的肝脏、eWAT和scWAT中均无显著变化(图4L和4M),研究表明肝脏FGF21可调控肝脏脂肪酸氧化相关基因(Pparα、Cpt1a、Pgc1a)和胆汁酸合成相关基因(Cyp7a1)的表达。结果显示,与对照小鼠相比,TG小鼠肝脏中仅Pgc1a基因表达上调(图4K和4N)。
先前研究表明,肝脏FGF21过表达可导致生长迟缓并诱导酮体生成。而在HFD-Dox饲喂1.5年的衰老TG小鼠中发现血清总酮体水平低于对照小鼠(图4O),这与肝脏中Pck1的基因表达升高(Pck1编码磷酸烯醇式丙酮酸羧化激酶PEPCK,该酶活性升高会降低脂肪酸氧化进而减少酮体生成)和酮体生成相关基因Hmgcs表达下降一致(图4P)(小编注:脂肪组织过表达FGF21小鼠的肝脏FGF21 mRNA和蛋白表达水平下降,且肝脏中FGF21受体Fgfr-1c基因表达下降,可能血清中脂肪来源FGF21含量长期升高,减少了对肝脏FGF21表达的需求以及肝脏对FGF21的响应,从而导致脂肪过表达FGF21小鼠与肝脏过表达FGF21小鼠的差异)。
总之,以上结果表明肝脏和脂肪组织中过表达FGF21会对小鼠肝脏代谢产生不同的影响,在HFD-Dox饲喂的衰老小鼠模型中,脂肪组织过表达FGF21通过降低肝脏脂质异位沉积而改善肝脏功能。
图S4.脂肪细胞特异性过表达FGF21可维持白色脂肪组织的脂质代谢,并防止高脂饮食小鼠在饲养1.5年后出现肝脂肪变性
9.过表达FGF21不影响HFD小鼠冷诱导的产热作用。
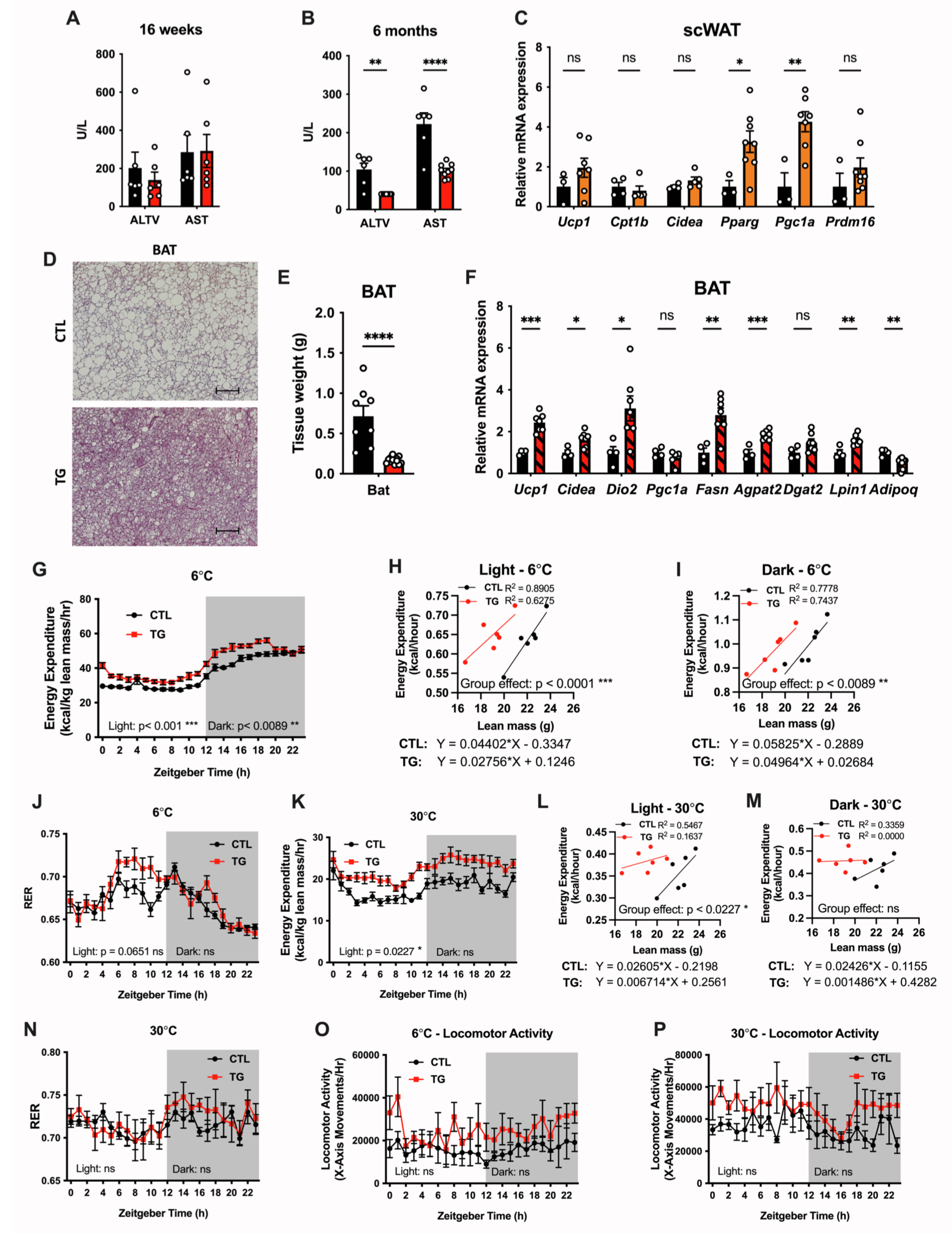
先前研究报道,肝脏分泌的FGF21激活了BAT和scWAT中的产热脂肪细胞,进而改善了心脏代谢健康。因此,研究人员想要探究脂肪组织特异性过表达FGF21对脂肪组织产热的影响。结果显示HFD-Dox饲喂1.5年的TG小鼠scWAT中UCP1基因表达无明显变化,但PPARγ和Ppargc1a基因表达显著上调;BAT中产热相关基因Ucp1、Cidea和Dio2表达显著上调,且脂肪生成相关基因Fasn、Agpat2和Dgat2表达显著上调(图S4C,S4F),然而,TG小鼠BAT中脂联素基因表达降低(图3K),H&E染色结果显示,TG小鼠的BAT发生显著去脂化,且BAT重量减轻(图S4D和S4E)。此外,TG小鼠的scWAT脂肪细胞更小,但没有发生棕色化(图3I)。
由于环境温度会显著影响棕色或米色脂肪细胞的产热作用,为了评估脂肪组织过表达FGF21是否能提高小鼠耐寒性,研究人员将HFD-Dox饲喂10周的对照小鼠和TG小鼠置于6℃环境下24h,监测机体能量消耗,结果显示以Lean mass归一化计算,TG小鼠的能量消耗显著升高,而ANCOVA分析结果显示在对照小鼠与TG小鼠之间Lean mass对机体能量消耗的影响相似(图S4G-S4I)。在22°C环境下,TG小鼠的RER没有差异(图2O)。然而,在6°C环境下,TG小鼠在光照阶段的RER高于对照小鼠,这表明脂肪组织FGF21促进机体利用碳水化合物供能(小编注:先前研究表明肝脏过表达FGF21可促进肝脏脂肪酸氧化途径,而在本文中,研究结果发现脂肪组织过表达FGF21促进了eWAT和scWAT中脂生成途径,抑制了脂解途径,没有检测eWAT和scWAT中脂肪酸氧化途径的变化,且脂肪组织过表达FGF21对肝脏脂肪酸氧化途径无显著影响。而研究发现TG小鼠eWAT和scWAT中胰岛素信号途径增强,促进葡萄糖摄取和利用,这可能促进了小鼠在冷刺激下的碳水化合物的利用),以响应寒冷反应(图S4J)。随后,研究人员监测了HFD-Dox饲喂10周的对照小鼠和TG小鼠在热中性(30℃)环境中的机体能量消耗,发现在光照阶段,TG小鼠能量消耗显著增加,RER没有差异(图S4K-S4N)。TG小鼠在30°C或6°C环境下的活动量均没有差异(图S4O和S4P)。总之,这些结果表明,尽管脂肪组织过表达FGF21促进了scWAT和BAT中产热相关基因的表达,但并不影响机体能量消耗。而在热中性环境下TG小鼠的能量消耗显著增加,这表明脂肪组织FGF21对能量消耗的影响并不依赖于BAT的功能,即使BAT组织中高表达FGF21。
10.过表达FGF21可缓解脂肪组织炎症
在衰老和肥胖进程中,脂肪组织发挥重要免疫活性,能进行先天和适应性免疫反应。脂肪细胞或浸润的巨噬细胞会释放细胞因子驱动慢性低度炎症,导致胰岛素抵抗以及与肥胖和衰老相关的疾病。瘦素是一种促炎性脂肪因子,通常在肥胖情况下含量增加,导致慢性低度炎症。研究人员发现,与HFD-Dox饲喂1.5年的对照小鼠相比,TG小鼠血清瘦素水平显著下降,脾脏重量减轻(图5A-5B),这提示TG小鼠机体炎症水平下降。
对HFD-Dox饲喂1.5年的衰老对照小鼠和TG小鼠eWAT组织蛋白质组学中表达下调的蛋白进行GO分析,发现与免疫反应通路相关的蛋白质显著减少(图5C和S5A)。而scWAT组织蛋白质组学中表达下调的蛋白在生物学过程(GO_bp)分析中仅有一条通路发生变化,在分子功能(GO_mf)分析无通路发生变化,这表明FGF21对脂肪组织免疫功能的调节具有组织特异性(图5D)。巨噬细胞是肥胖期间脂肪组织中最丰富的免疫细胞类型,能引发并促进炎症反应。与HFD-Dox饲喂1.5年的衰老对照小鼠相比,TG小鼠eWAT中的巨噬细胞总数没有变化(图5E),但巨噬细胞亚型发生显著转变,M1促炎性巨噬细胞数量减少,而M2抗炎性巨噬细胞数量增加,表明脂肪组织过表达FGF21可减轻炎症反应(图5F–5I)。
衰老与效应记忆细胞的异常扩增、天然T淋巴细胞的丧失以及脂肪B细胞的增加有关。FGF21可通过改善胸腺上皮细胞的功能来保护胸腺免于退化,这对于维持淋巴细胞发育至关重要。研究人员发现,与HFD-Dox饲喂1.5年的衰老小鼠相比,TG小鼠eWAT中B细胞数量下降,CD8细胞数量下降,CD4细胞数量无明显变化(图5J,5N-5Q)。这与先前的研究结果一致,即耗竭脂肪组织B细胞可以改善衰老小鼠胰岛素敏感性(小编注:研究报道,在衰老小鼠eWAT中会积累一种非衰老性衰老脂肪B细胞(AABs),主要在脂肪相关淋巴簇(FALCs)中积累,通过依赖Nlrp3炎症小体和IL-1信号途径损害脂肪细胞的代谢功能。向eWAT注射CD20mAb以耗竭B细胞可改善衰老小鼠的胰岛素敏感性)。同时,TG小鼠eWAT中NK细胞数量没有差异(图S5B)。研究报道饮食诱导的肥胖通常伴有γδT细胞的增加,结果显示TG小鼠eWAT中促炎性CD44+γδT细胞数量增加,总γδT细胞数量无变化(图5K-5L)。此外,TG小鼠eWAT中Tregs(调节性T细胞)数量和ILC2s(2型先天淋巴细胞)显著增加(小编注:研究发现,衰老小鼠VAT中ILC2s水平降低。研究人员对衰老小鼠腹腔注射外源性IL-33(维持ILC2s的主要调节因子),提高了衰老小鼠VAT中的ILC2s数量,但并没有改变衰老小鼠的能量消耗和糖稳态,甚至提高了冷刺激下小鼠的死亡率,机制上,补充IL33提高的ILC2s具有衰老、促炎特征,进而损害衰老小鼠的代谢。另外,有研究表明,在衰老过程中小鼠VAT组织中Treg细胞数量增加,消耗VAT中Treg细胞可改善衰老小鼠胰岛素敏感性;然而,在肥胖状态下VAT中Treg水平降低,这一差异与不同Treg亚型有关,CD73high亚型具有抗炎表型,可改善胰岛素敏感性,而ST2high亚型抑制米色脂肪生成,具有促炎表型,主要在衰老中富集。本篇文章中,研究人员发现在长期HFD喂养下,衰老TG小鼠eWAT中ILC2s数量增加,这逆转了衰老相关的ILC2s数量减少现象;此外,TG小鼠eWAT中Treg细胞数量增多(文章没有描述具体的Treg亚型),这提示FGF21在eWAT中可能发挥抗炎调节作用)(图5M,5R),总之,这些结果表明FGF21可维持脂肪组织内驻留免疫细胞的稳态,抑制“炎症性衰老”。
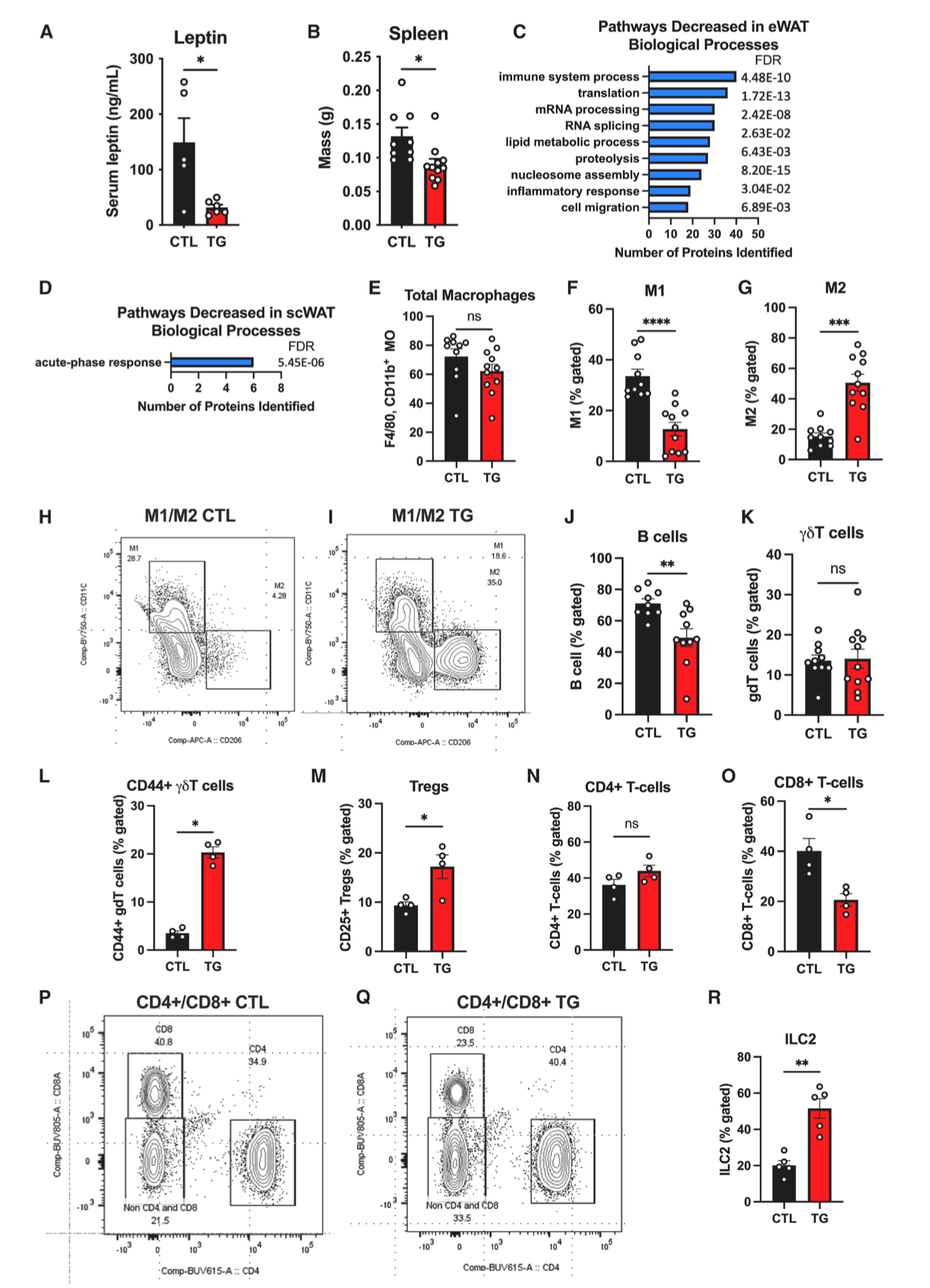
图5.FGF21过表达可改善高脂饮食衰老小鼠内脏脂肪组织的免疫代谢功能
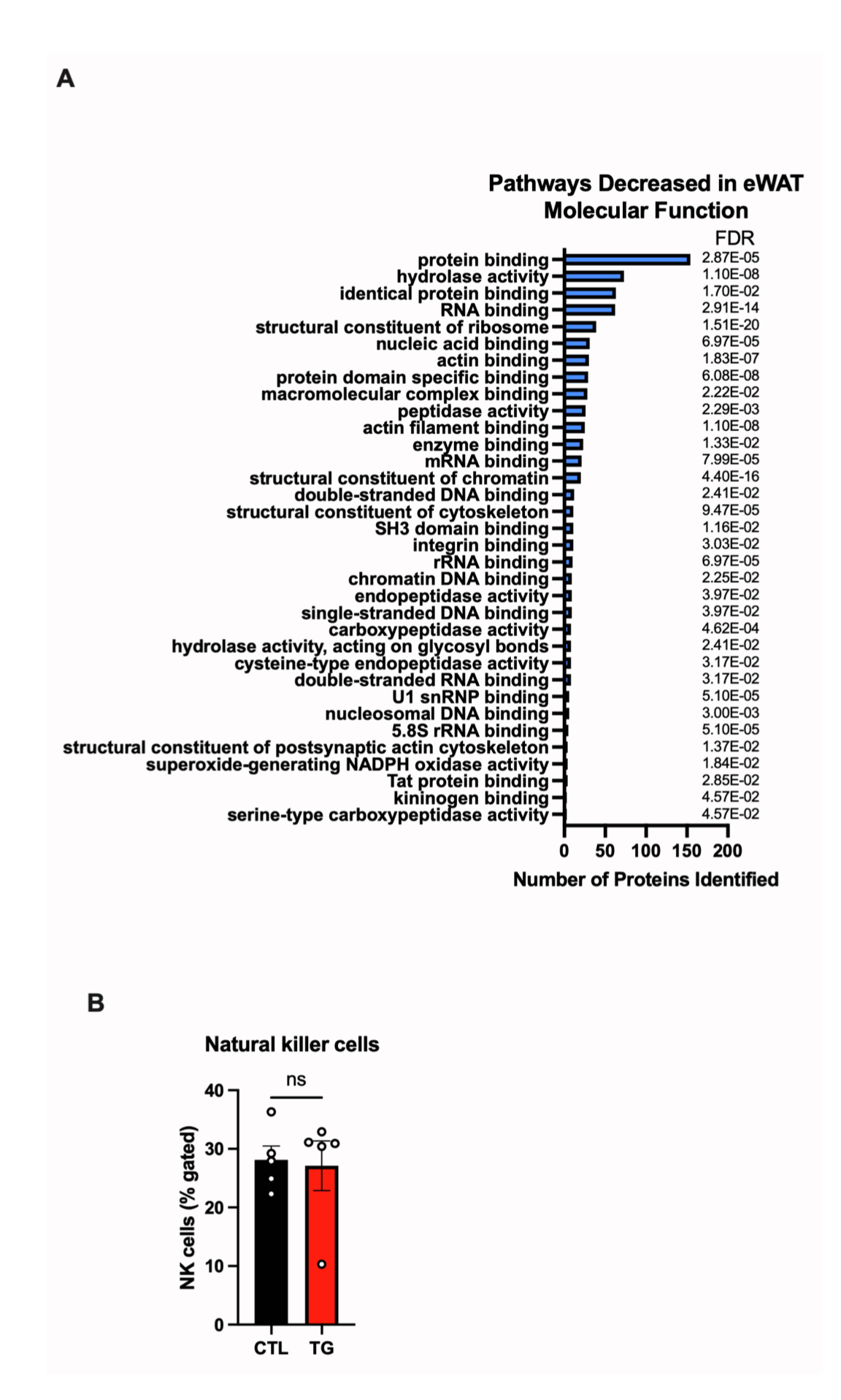
图S5.FGF21过表达可改善高脂饮食衰老小鼠内脏脂肪组织的免疫代谢功能
11.过表达FGF21可降低内脏脂肪组织中神经酰胺水平
接下来,研究人员探究了FGF21调控eWAT组织炎症反应的机制。由于FGF21的受体βKlotho在巨噬细胞或T细胞中不表达,这表明FGF21可能是通过eWAT中细胞间交流来介导免疫反应。神经酰胺是一种脂质代谢物,肥胖期间在代谢组织中含量增加,通过抑制胰岛素信号传导等机制引起细胞功能障碍。有研究表明FGF21可降低神经酰胺水平,但机制仍不清楚。因此,研究人员对HFD-Dox饲喂1.5年的衰老对照小鼠和TG小鼠进行液相色谱-串联质谱(LC-MS/MS)分析,发现TG小鼠eWAT中几种神经酰胺水平(C16、C18、C20、C22和C24:1)显著减少(图6B)。而TG小鼠scWAT中神经酰胺C24和C25水平升高(图6C)。基因表达分析显示,TG小鼠eWAT中神经酰胺合酶基因Cer5、Cer6和Degs表达降低,而scWAT中Cer2和Sptlc1表达上调(图6D和6E)。此外,TG小鼠eWAT中调节神经酰胺代谢的蛋白质发生了显著变化,其中3-酮二氢鞘氨醇还原酶(KDSR)表达增加,神经酰胺合酶2(CERS2)、神经酰胺转运蛋白1(CERT1)、Δ4去饱和酶(DEGS)、鞘氨醇-1-磷酸裂解酶(SGPL1)和N-酰基鞘氨醇酰胺水解酶1(ASAH1)表达减少(图6F)。而TG小鼠scWAT中参与神经酰胺代谢的蛋白质表达水平没有变化。
神经酰胺生物合成的第一步是棕榈酰基辅酶A(CoA)和丝氨酸通过丝氨酸棕榈酰基转移酶进行缩合反应,产生鞘氨醇(二氢鞘氨醇)(图6A)。与HFD-Dox饲喂1.5年的衰老对照小鼠相比,TG小鼠eWAT中丝氨酸和鞘氨醇水平下降,而scWAT中无明显变化(图6G-6I)。此外,TG小鼠eWAT和scWAT中二氢神经酰胺中间体的水平无显著差异(图S6A和S6B)。
神经酰胺还可通过鞘氨醇或鞘磷脂途径合成或分解代谢。结果显示,与HFD-Dox饲喂1.5年的衰老对照小鼠相比,TG小鼠eWAT中鞘氨醇水平降低,而scWAT中无明显变化(图6H和6I)。总之,这些结果表明,脂肪组织过表达FGF21可降低eWAT组织中神经酰胺水平,而对scWAT中神经酰胺水平无影响。
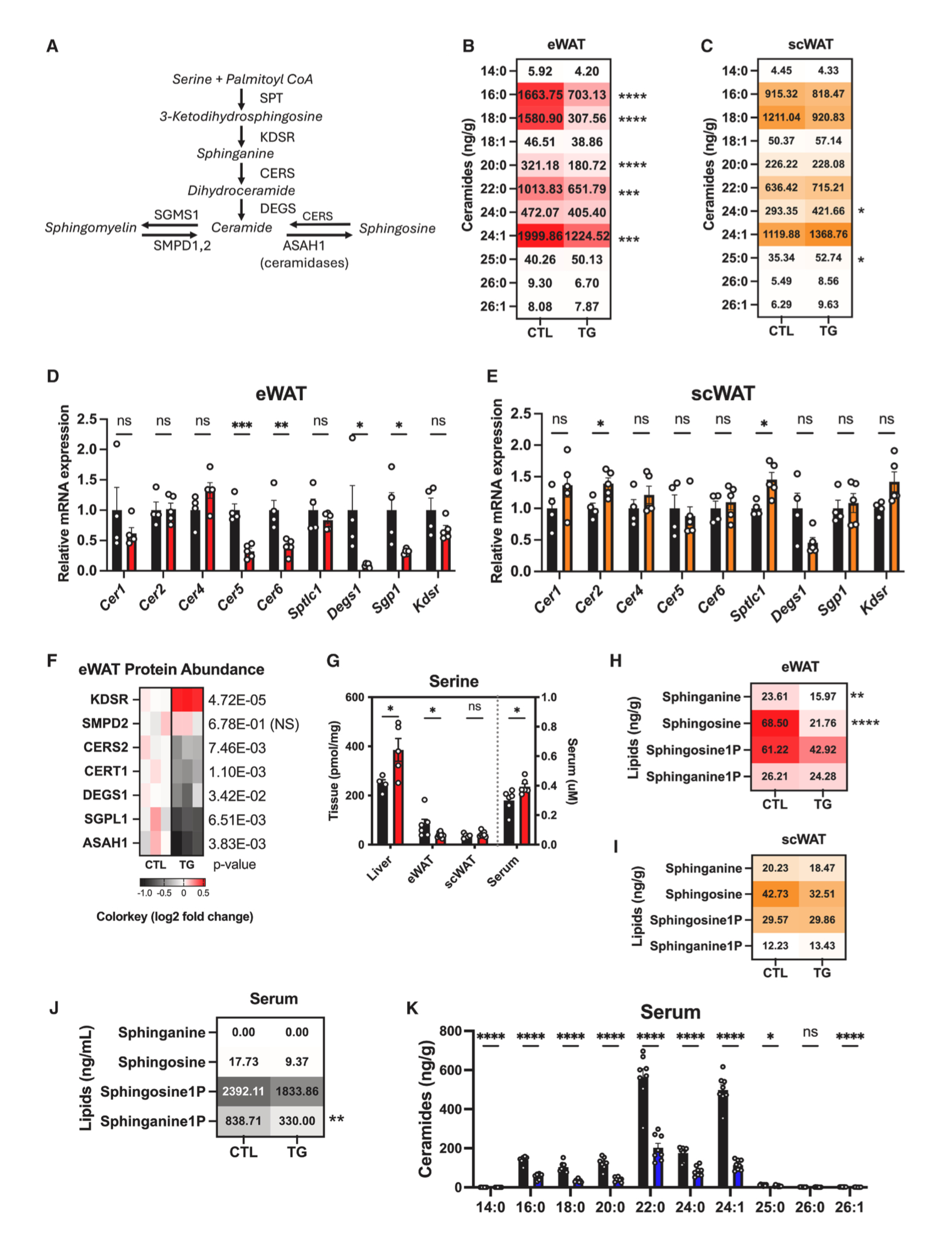
图6.脂肪细胞特异性FGF21过表达可降低高脂饮食衰老小鼠内脏脂肪组织及循环中的鞘脂和神经酰胺水平
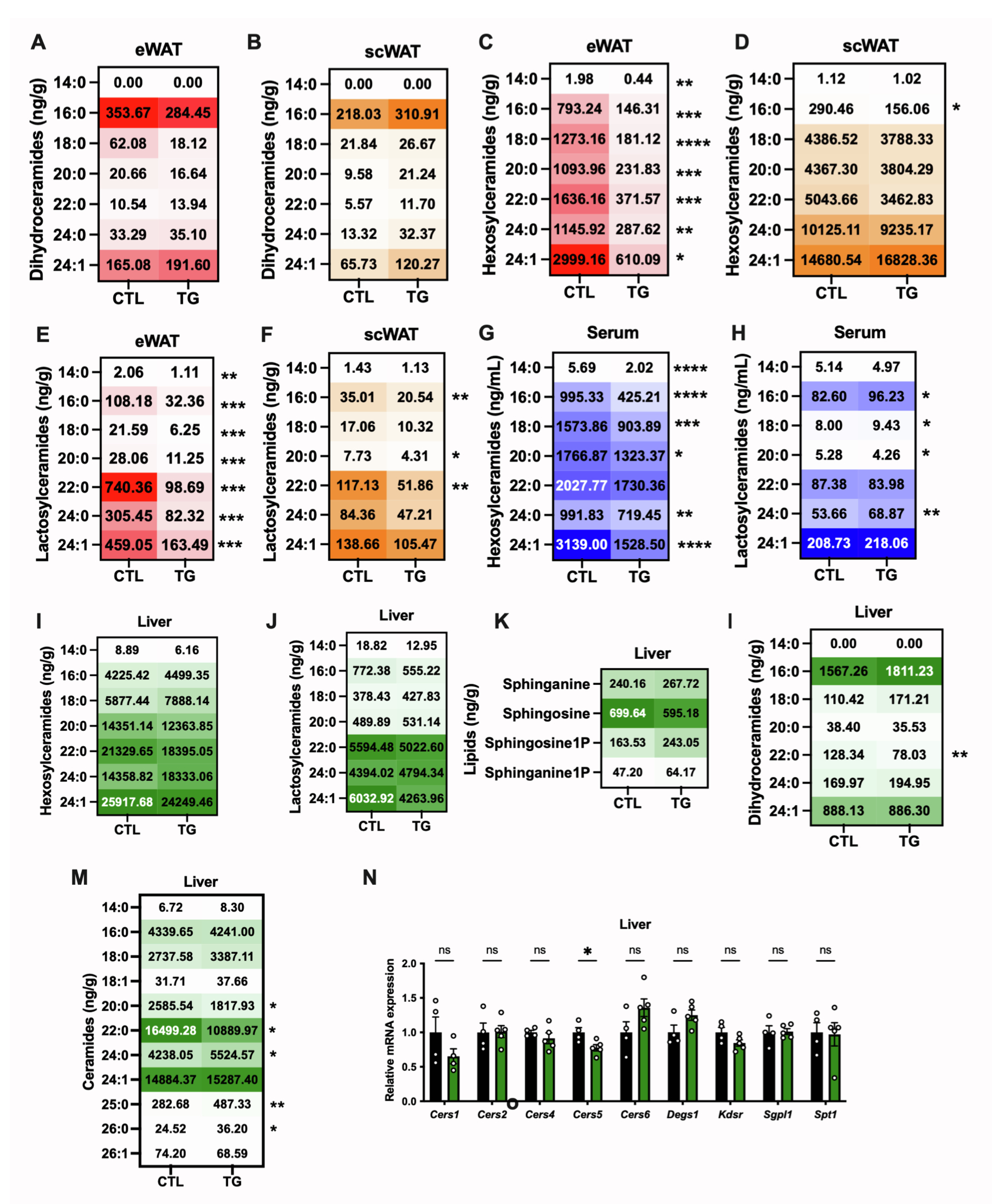
图S6.脂肪细胞特异性FGF21过表达可降低高脂饮食衰老小鼠内脏脂肪组织及循环中的鞘脂和神经酰胺水平
12.过表达FGF21可降低脂肪组织中的鞘脂水平
神经酰胺是合成复杂鞘糖脂(如己糖基神经酰胺和乳糖基神经酰胺)的前体。在衰老过程中,这些具有生物活性的鞘脂会在多种器官中积累,并与多种疾病相关。结果显示,与HFD-Dox饲喂1.5年的衰老对照小鼠相比,TG小鼠eWAT中己糖基神经酰胺和乳糖基神经酰胺(包括C14、C16、C18、C20、C24和C24:1等种类)含量显著减少(图S6C和S6E),scWAT中己糖基神经酰胺C16以及乳糖基神经酰胺C16、C20和C22水平同样降低(图S6D和S6F)。此外,TG小鼠血清中多种己糖基和乳糖基神经酰胺含量降低(图S6G和S6H)。这些鞘脂与氧化应激和炎症反应密切相关,因此研究人员认为eWAT中神经酰胺的减少可能是FGF21发挥抗炎效应的机制,但FGF21调控这些鞘脂水平的具体机制仍需进一步研究。
13.脂肪组织FGF21过表达可降低肥胖衰老小鼠循环神经酰胺水平
循环神经酰胺水平升高与衰老、心血管疾病、中风和2型糖尿病的高发病率密切相关。研究人员进一步检测了血清中与心血管疾病和胰岛素抵抗相关的神经酰胺种类(如C16、C18、C18:1、C24和C24:1)的含量,探究其含量的变化是否受脂肪组织FGF21的影响。结果发现,与HFD-Dox饲喂1.5年的衰老对照小鼠相比,TG小鼠血清中多种神经酰胺(包括C14、C16、C18、C20、C22、C24、C24:1、C25和C26:1)以及鞘氨醇-1-磷酸的水平显著降低(图6J和6K)。
肝脏中含有高水平的神经酰胺,肝脏中神经酰胺的变化可影响到全身神经酰胺的代谢和分布。结果显示,与HFD-Dox饲喂1.5年的衰老对照小鼠相比,TG小鼠肝脏鞘氨醇和鞘氨烷水平未发生明显变化,且己糖基神经酰胺与乳糖基神经酰胺含量也没有明显变化(图S6I-S6K),而TG小鼠肝脏中的神经酰胺(C20、C22)和二氢神经酰胺(C22)水平显著减少,这与血清中观察到的下降趋势一致(图S6L和S6M)。TG小鼠肝脏中部分长链神经酰胺(如C24、C25和C26)显著增加(图S6M)。神经酰胺代谢相关基因表达结果显示,TG小鼠肝脏中仅神经酰胺合成酶5(Cers5)的表达下调(图S6N)。总之,这些结果表明,不同于以往的血清神经酰胺水平主要受肝脏神经酰胺合成的影响,在HFD-Dox饲喂的衰老TG小鼠模型中,脂肪组织多种神经酰胺的减少介导了血清中多种神经酰胺含量的下降,而这种变化可能与减轻的炎症反应和增强的胰岛素敏感性相关。
14.FGF21独立于脂联素的作用来降低内脏脂肪中神经酰胺含量
前面数据发现脂肪组织过表达FGF21显著增加了血清脂联素水平(图3L),脂联素作为强效抗炎因子,其受体具有内在的鞘氨醇酶活性,能够促进神经酰胺的分解。且先前已有研究表明FGF21对脂肪组织功能稳态的调控是通过脂联素介导的,因此研究人员构建了脂联素敲除小鼠(ADNKO小鼠),并与脂联素启动子驱动的rtTA小鼠和TRE-FGF21小鼠杂交,产生ADNKO小鼠和脂肪组织过表达FGF21的ADNKO小鼠,所有小鼠均饲喂HFD-Dox饮食持续1.5年。结果显示,与衰老WT小鼠相比,ADNKO小鼠eWAT和scWAT中鞘脂水平升高,而过表达FGF21后显著降低了eWAT中鞘脂,scWAT中鞘脂水平无明显变化(图7A-7D、S7A-S7B)。此外,与衰老ADNKO小鼠相比,过表达FGF21显著降低了eWAT多种神经酰胺的含量,而scWAT中神经酰胺水平有所增加 (图7E和7F)。这表明eWAT对FGF21信号更加敏感,FGF21调控eWAT中的神经酰胺水平不需要依赖于脂联素。相比之下,scWAT对脂联素信号更加敏感,在脂联素缺乏的情况下,即使过表达FGF21,scWAT中神经酰胺水平仍会增加,这表明FGF21和脂联素在eWAT和scWAT中的作用存在组织特异性。
总体而言,FGF21在eWAT中的信号传导是脂肪组织调控机体肥胖和衰老的一个重要因素,并强调了脂肪组织作为代谢紊乱治疗靶点的潜力。在衰老小鼠中FGF21对能量消耗无明显作用,这表明FGF21可能通过其他方面发挥延长寿命的作用,这可能与FGF21降低神经酰胺水平的作用相关。
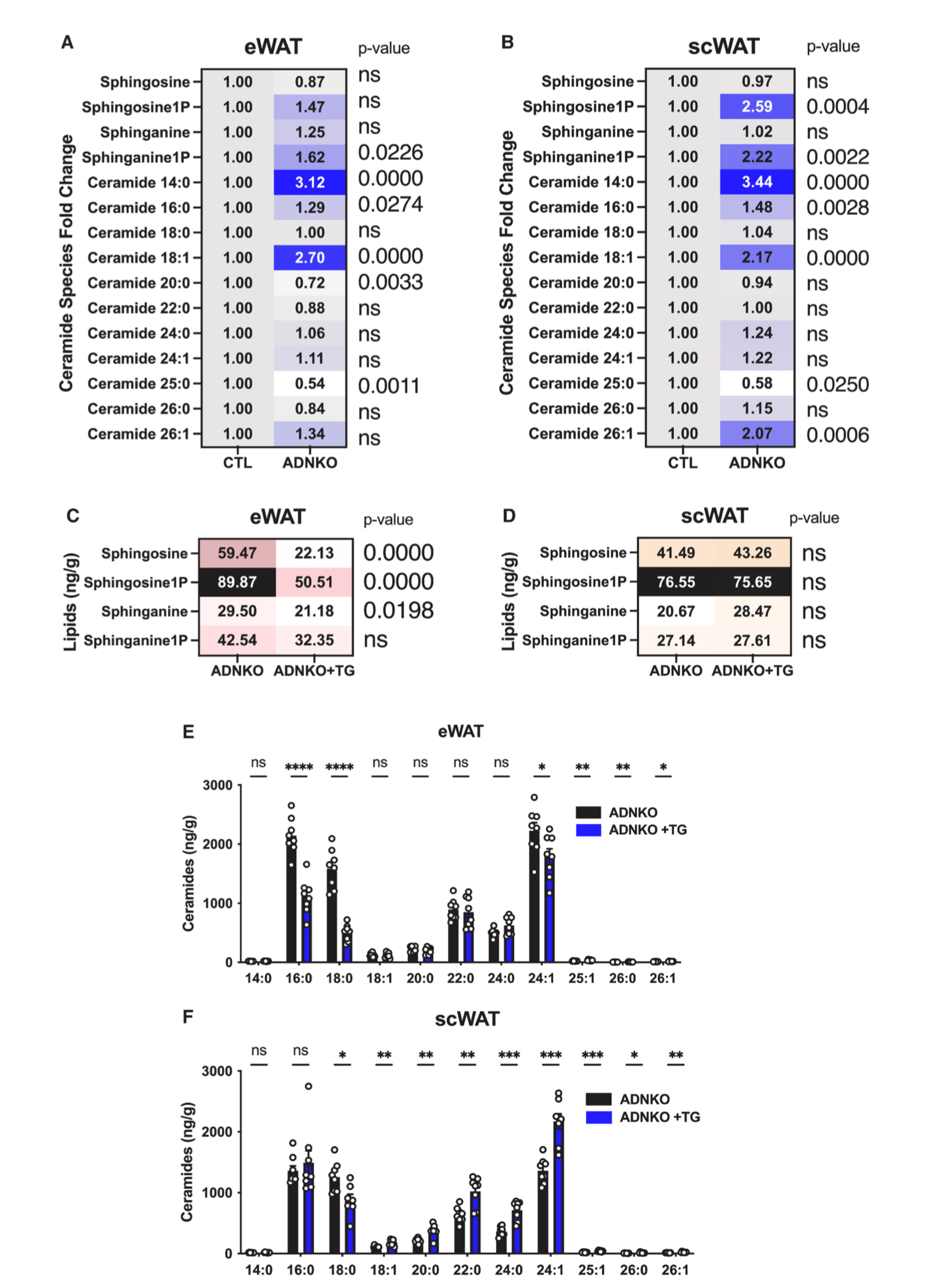
图7.FGF21过表达可逆转ADNKO小鼠内脏脂肪组织中神经酰胺的升高
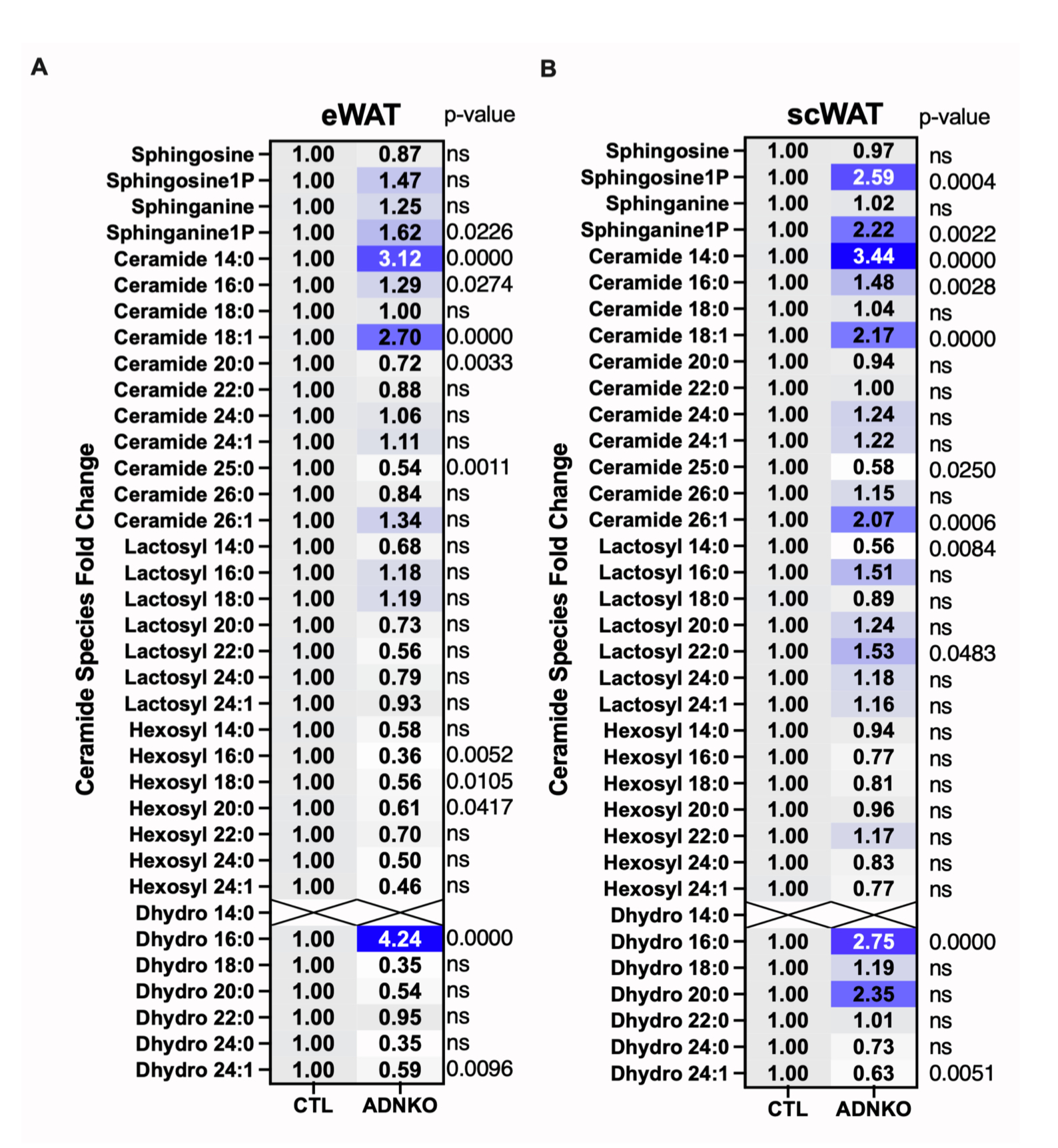
图S7.FGF21过表达可逆转ADNKO小鼠内脏脂肪组织中神经酰胺的升高
总结
本篇文章研究了脂肪细胞特异性过表达FGF21对HFD诱导的肥胖小鼠寿命和代谢健康的促进作用。研究发现,成年小鼠脂肪细胞特异性过表达FGF21能显著延长寿命并改善代谢健康,包括减少体重增加、提高胰岛素敏感性、降低肝脏脂肪变性和内脏脂肪炎症。并且这些益处与生长抑制无关,且不依赖于能量消耗的增加。进一步研究发现,FGF21通过降低内脏脂肪中的神经酰胺水平,减少炎症性免疫细胞浸润,从而改善代谢功能。此外,FGF21的作用部分独立于脂肪激素脂联素,提示其通过多种机制发挥作用。这些结果揭示了FGF21在改善肥胖相关代谢紊乱和延长健康寿命中的潜在作用,为治疗代谢综合征和年龄相关疾病提供了新的治疗靶点。
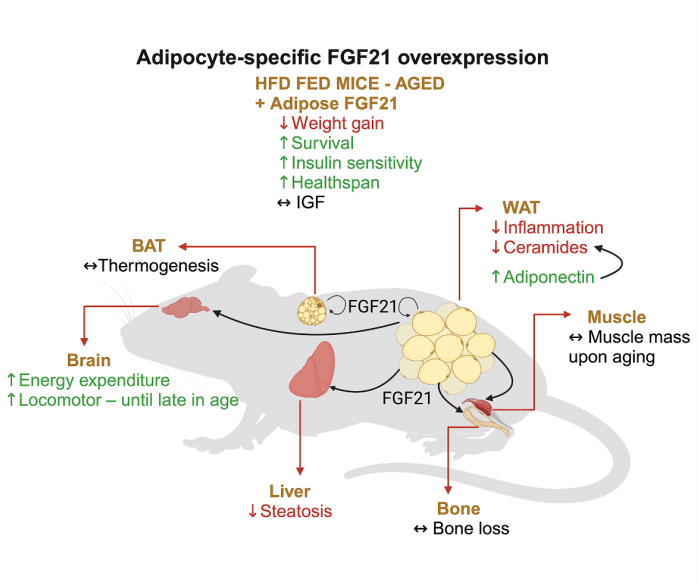
原文链接:https://www.sciencedirect.com/science/article/abs/pii/S1550413125002670?via%3Dihub
转载本文请联系原作者获取授权,同时请注明本文来自徐凌燕科学网博客。
链接地址:https://wap.sciencenet.cn/blog-3483272-1505257.html?mobile=1
收藏