博文
植物的联想学习:研究历史、核心争议与进展  精选
精选
|
![]()
核心概览:“联想学习”作为一种高级适应性行为,长期被认为是动物界的专属特征,其核心是有机体通过后天经验建立两个刺激间的关联并调整行为。从20世纪60年代的初步探索到2016年的突破性报道,植物联想学习的研究历经半个多世纪的起伏。2016年Gagliano团队首次通过Y型迷宫实验证实豌豆 可将风扇气流与蓝光建立关联,颠覆传统认知;2020年Markel的重复实验未能重现该结果,引发激烈争议,同年《eLife》同期刊登双方回应——Gagliano团队指出光照环境控制不足导致非条件刺激失效,Markel则通过直方图匹配技术论证实验设计差异无法解释阴性结果;2022年Cvrčková与Konrádová的研究进一步提出前期光照质量历史是关键混淆变量。本文系统梳理植物联想学习的研究脉络、核心争议与最新进展,深入剖析争议背后的科学逻辑与方法论差异,旨在全面呈现该领域的研究价值与未来方向。

1 植物联想学习的研究历史与科学意义
1.1 研究历史:从初步探索到突破性发现
植物联想学习(associative learning)的研究可追溯至20世纪60年代,早期学者已尝试探索植物是否具备类似动物的条件反射能力:
奠基阶段(1960s-1970s):Holmes与Gruenberg(1965)首次提出“植物可能存在学习”的假设,通过对豌豆(Pisum sativum)幼苗的简单刺激实验初步探索条件反射[13];Haney(1969)与Levy等人(1970)分别以含羞草(Mimosa pudica)为研究对象,尝试通过机械刺激与光照的配对训练诱导条件反射,但实验设计缺乏严格对照,结果难以排除非特异性反应的干扰[7];Armus(1970)的研究虽声称观察到含羞草的条件反射,但因未控制环境变量被后续研究质疑[7]。这一阶段的研究因方法学局限,未能提供植物联想学习的可靠证据,相关探索陷入停滞。
复苏阶段(2010s):随着植物行为学与分子生物学的发展,研究者开始重新审视植物的认知能力。澳大利亚西澳大学Gagliano团队(2014)发现植物可通过习惯化(habituation)实现非联想学习,为联想学习的探索奠定基础[1];2016年,该团队在《Scientific Reports》发表开创性研究,通过严谨的Y型迷宫实验首次报道豌豆的联想学习能力,引发学界广泛关注[1]。
争议与深化阶段(2020s至今):2020年美国加州大学戴维斯分校 Markel的重复实验未能重现Gagliano团队的结果,《eLife》同期刊登双方学术回应,聚焦实验设计差异的核心争议[3,4];2022年捷克查理大学 Cvrčková与Konrádová提出光照质量历史的全新视角,推动研究向更精细的环境变量控制方向发展[6];同时,多组学技术的应用为解析植物联想学习的分子机制提供了新工具。
1.2 科学意义:重塑对学习与生命适应的认知
进化生物学价值:若植物确实存在联想学习,将打破“联想学习仅存在于后生动物(metazoa)”的传统认知,证明该能力是动植物共有的通用适应性机制[1]。这一发现将重塑学习行为的进化史,为探索认知能力的起源与趋同进化提供关键线索[1,4]。
植物行为学突破:传统观点认为植物行为多为先天本能,而联想学习的证实意味着植物具备基于经验调整行为的灵活性,推动植物行为学从“本能应答研究”向“认知调控探索”转型[1,6]。
跨学科研究推动:该领域的研究融合了植物生理学、神经科学、进化生物学与哲学,引发对“学习” “记忆” “认知”等核心概念的重新界定,促进跨学科理论与方法的交叉融合[1,7]。
2 争议的起点:植物联想学习的首次证实(2016)
2.1 实验设计与核心发现
澳大利亚西澳大学 Monica Gagliano团队借鉴巴甫洛夫经典条件反射范式,以豌豆(Pisum sativum cv Massey Gem) 为研究对象开展实验[1]。实验中,蓝光(blue LED light) 作为非条件刺激(unconditioned stimulus, US)——利用植物先天的向光性(phototropism)确保稳定应答;35mm直流风扇产生的气流(wind from DC12v mini fan) 作为条件刺激(conditioned stimulus, CS)——预实验证实气流本身对豌豆生长方向无显著影响,属于中性刺激。
实验设置两组核心处理:F+L组(风扇与蓝光位于Y型迷宫同一臂)和F vs L组(风扇与蓝光位于迷宫对侧臂)。经过3天训练(每天3个训练时段,风扇提前蓝光60分钟启动,重叠30分钟)后,测试阶段仅保留气流刺激。结果显示:对照组(无风扇刺激)豌豆100%向最后一次蓝光照射方向生长;测试组中,62%的F+L组豌豆向气流方向生长,69%的F vs L组豌豆向气流对侧生长,且两组均与对照组存在显著统计学差异(F+L组p=0.0027,F vs L组p=0.0017)[1]。
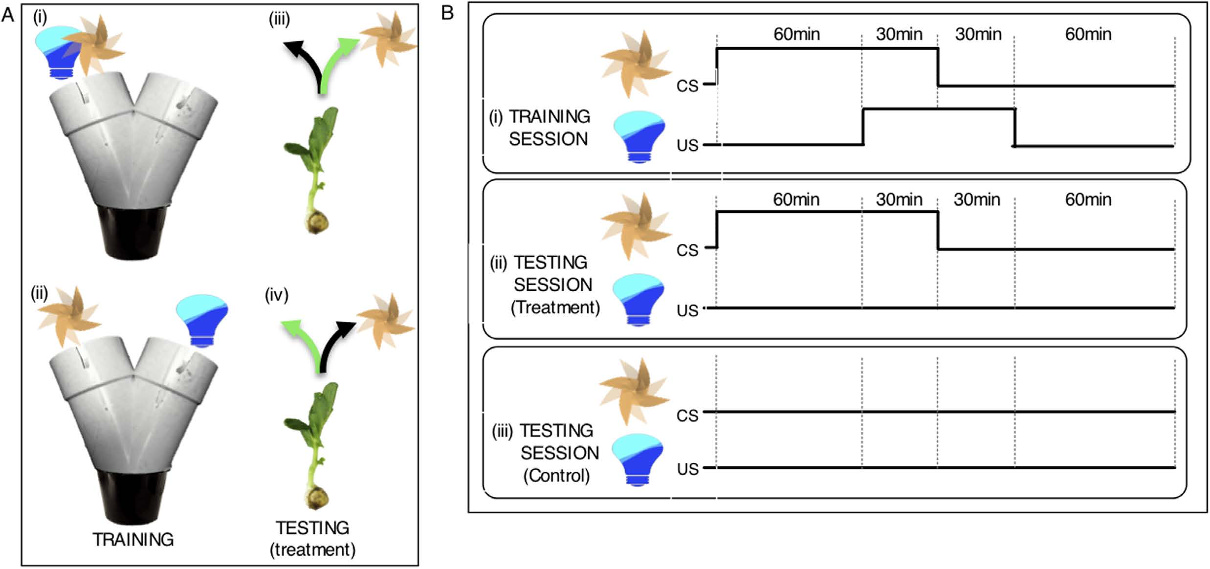
图1 豌豆联想学习的训练与测试方案
左图为训练阶段设计,F+L组(上)中风扇与蓝光位于迷宫同一臂,F vs L组(下)中二者位于对侧臂;右图为测试阶段设计,仅保留风扇刺激,箭头指示豌豆的条件应答方向(绿色为正确关联应答,黑色为未形成关联应答)。右图 幼苗在测试前接受连续三天训练,每天包含三个2小时训练 session,间隔1小时。90分钟的CS比60分钟的US提前60分钟呈现,重叠30分钟(i)。测试日,幼苗仅暴露于风扇三个90分钟 (ii);对照组幼苗不受干扰(无风扇、无光;iii)。Gagliano, M., et al. (2016)[1]
进一步的昼夜节律(circadian rhythm)实验发现,该联想学习具有时间特异性:仅在植物的“主观白天”(subjective day)训练才能成功,夜间或明暗交替阶段训练则无此效果,表明学习行为受生物钟调控[1]。Gagliano团队据此提出核心结论:联想学习并非动物专属,而是动植物共有的通用适应性机制[1]。
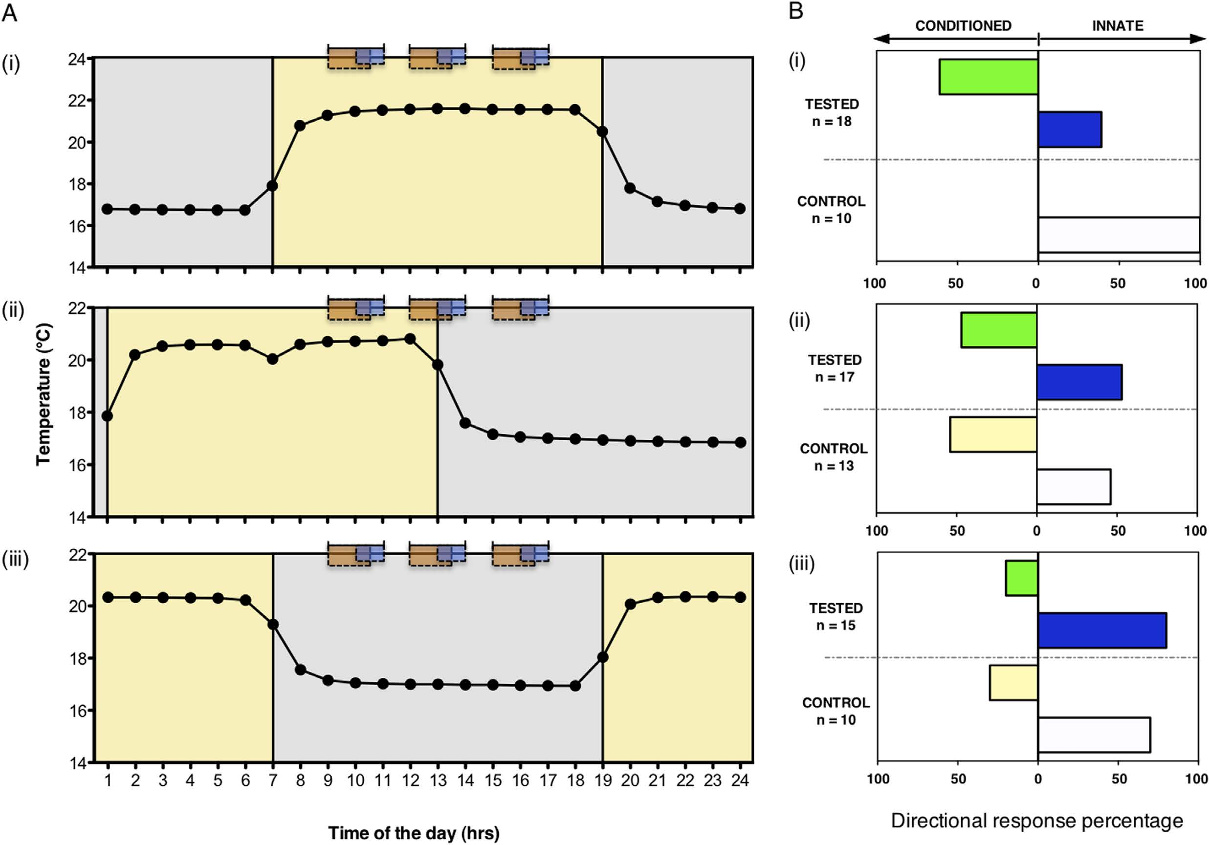
图2 昼夜节律对豌豆联想学习的影响
左图为三组实验的昼夜节律设置(Light组、Light-Dark组、Dark组),橙色与蓝色矩形分别表示风扇与蓝光刺激时段;右图为各组豌豆的生长应答比例,绿色柱代表形成联想学习的植株比例,白色柱代表保持先天向光性的植株比例,可见仅Light组存在显著的联想学习效应。Gagliano, M., et al. (2016)[1]
3 争议的爆发:重复实验与《eLife》同期学术回应(2020年6月)
3.1 Markel的重复实验与阴性结果
美国加州大学戴维斯分校 Kasey Markel于2020年在《eLife》发表重复实验结果[2],在核心条件(光照强度14 μmol m⁻² s⁻¹、温湿度20℃/85%、训练流程)上重复原研究,但做出两项关键优化:一是扩大样本量(如F+L组从31株增加至61株),提高统计效力;二是采用完全双盲分析(fully blinded analysis),排除实验者主观偏差[2,3]。
由于无法获取原实验材料,Markel选用亲缘关系相近的豌豆品种(Pisum sativum cv Green Arrow)[2]。实验结果显示:对照组豌豆仅轻微偏向最后一次蓝光照射方向(F+L对照组p=0.0243,F vs L对照组无统计学显著性),未出现100%向光性应答;核心实验组与对照组间无显著差异(F+L组p=0.335,F vs L组p=0.387)[2],未能重现联想学习现象。
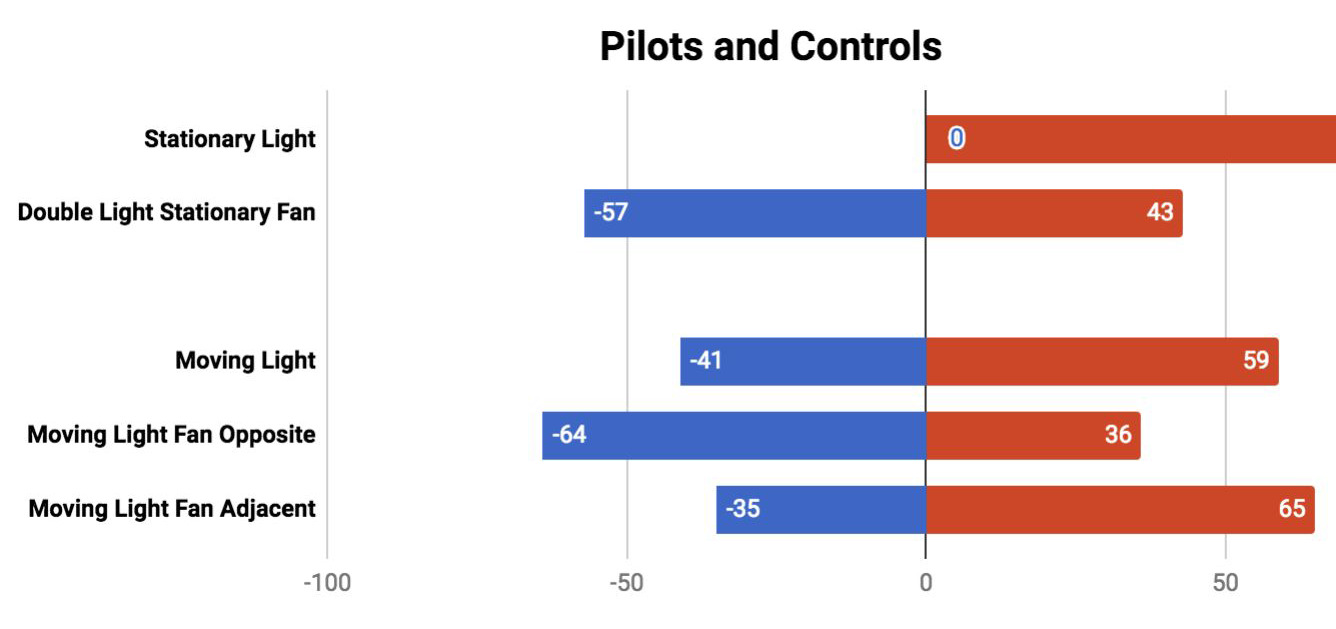
图3 原实验与重复实验的核心结果对比
Markel 研究(KM)与Gagliano等(2016)实验(MG)中植物在对照条件下的生长结果。柱状图中的数字表示百分比,括号内数字表示原始数据,即各迷宫臂中生长的植株数量。可见原实验中对照组100%向光生长,而重复实验中仅呈现轻微偏向,且实验组与对照组无显著差异。Markel, K. (2020)[2]
3.2 《eLife》同期登载的核心争议(2020年9月)
(1)Gagliano团队的反驳:实验设计缺陷导致非条件刺激失效[4]
Gagliano等人同期回应,指出Markel的实验方案不适用于验证联想学习,核心依据包括:
光照环境控制不足:原实验在5.3 m²全暗室中进行,豌豆间距约20 cm,确保光照仅来自目标LED;而Markel使用1.5 m²培养箱,植株间距紧凑,存在光源泄漏与反射,导致豌豆接收多源光照,干扰向光性应答,使蓝光无法成为有效US(非条件刺激需稳定引发应答,如巴甫洛夫实验中食物必然引发唾液分泌)。
昼夜节律的潜在影响:原实验已证实昼夜节律会影响蓝光的US有效性,黑暗阶段向光性减弱;虽Markel声称节律阶段一致,但培养环境差异可能导致相位偏差,进一步削弱向光性。
(2)Markel的回应:实验设计差异无法解释阴性结果[3]
Markel通过技术分析与实验数据反驳:
向光性应答的合理性辩护:多数植物向光性实验需3小时以上光照(Schumacher et al., 2018; Haga and Kimura, 2019),拟南芥黄化苗经1小时蓝光照射后向光性弯曲几乎未启动(Sullivan et al., 2016);且Y型迷宫中蓝光为斜向照射(与垂直方向夹角<40°),远弱于水平光照的诱导效果,原实验的100%向光性可能是特殊条件下的偶然结果。
光照环境差异的技术验证:采用直方图匹配技术分析显示,两组实验的背景亮度差异源于相机参数,而非实际光照强度(见图4);尽管植株间距更紧凑,但迷宫LED仍是主要光源,不足以解释向光性差异。
品种差异的无关性论证:选用的Green Arrow与Massey Gem亲缘关系近,均为全日照品种,联想学习作为复杂性状,不可能因短期品种分化而消失。
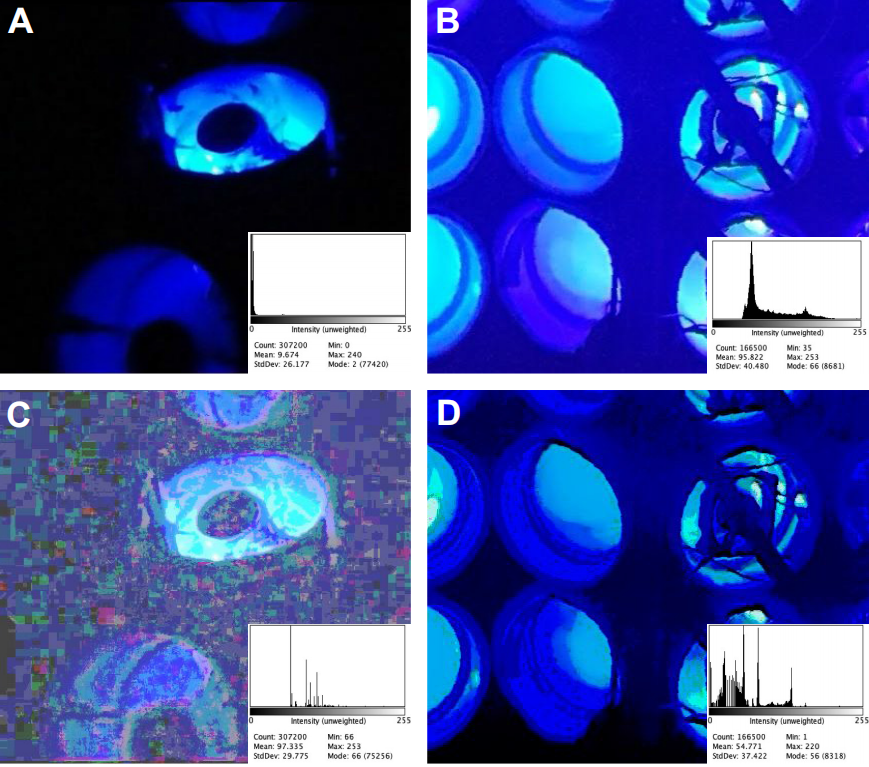
图4 原实验与重复实验的光照环境对比
左图为Gagliano 2016实验的Y迷宫光照场景(A)及直方图匹配后图像(C),右图为Markel 2020实验的光照场景(B)及直方图匹配后图像(D),底部直方图显示二者亮度差异主要源于相机参数,而非实际光照强度。Markel, K. (2020)[3]
4 争议的新维度与最新进展(2022年)
4.1 光照质量历史:被忽略的关键变量[6]
2022年,捷克查理大学 Fatima Cvrčková与Hana Konrádová在《BIOCELL》发表研究,提出双方此前均未关注的核心变量——实验前幼苗培养阶段的光照质量(light quality history)。
研究指出,Gagliano 2016与Markel 2020的前期培养光照光谱存在本质差异:Gagliano团队采用冷白色荧光灯(含绿色光等完整光谱),而Markel使用红蓝光LED组合(缺乏绿色光等关键波段)[6]。植物对光照质量高度敏感,光谱组成会通过表观遗传修饰等机制影响光受体系统(如光敏色素、向光素)的表达与活性,进而长期调控生长特性与环境应答能力[6]。例如,绿色光可诱导植物的避阴反应(Zhang et al., 2011),红蓝光比例会调控茎伸长与光合作用(Bayat et al., 2018)[6]。该研究推测,前期光照质量差异可能导致两组豌豆的光感知能力不同,最终使得Markel实验中蓝光无法稳定引发向光性应答,这一观点为争议提供了全新解读视角[6]。
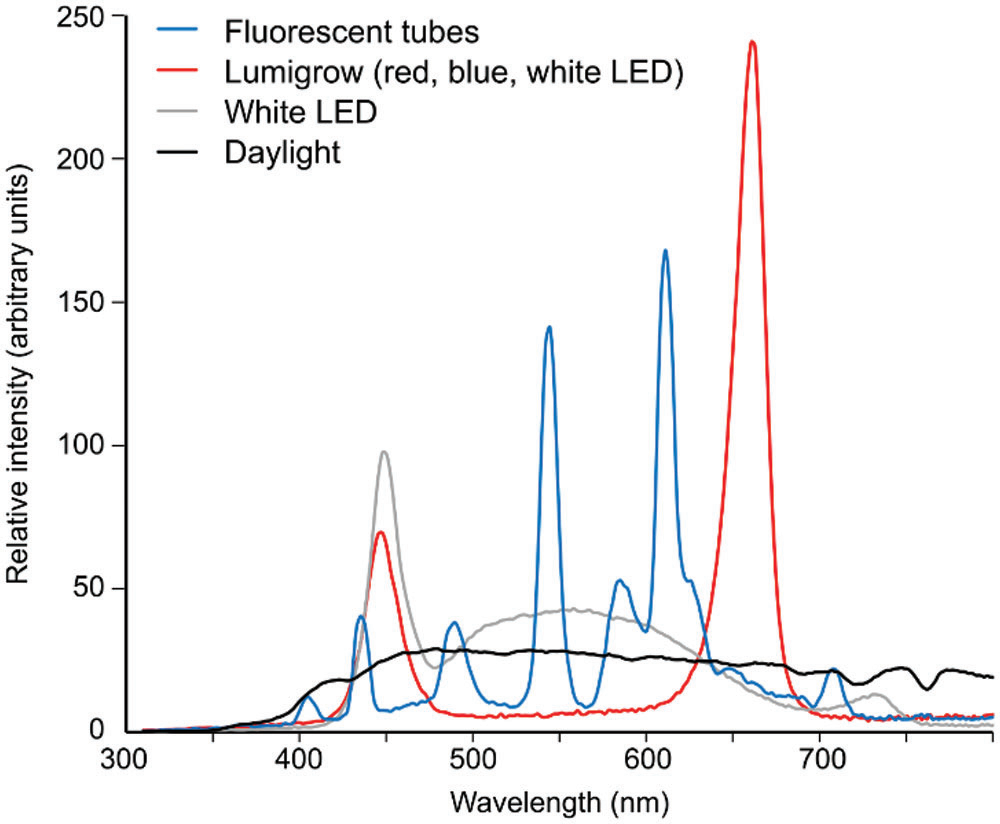
图5 几种常用于实验植物培养的光源的光谱特性示例
展示冷白色荧光灯(类似Gagliano 2016使用光源)、红蓝光LED组合(类似Markel 2020使用光源)、白色LED及自然 daylight 的光谱分布,可见荧光灯包含完整的可见光波段,而红蓝光LED组合缺乏绿色光等中间波段。Cvrčková, F., & Konrádová, H. (2022)[6]
4.2 分子机制探索的前沿进展
近年来,研究者开始尝试解析植物联想学习的潜在分子机制,虽尚未取得突破性成果,但已有多项研究提供线索:
表观遗传调控:Gagliano团队(2014)发现植物习惯化学习与表观遗传重编程相关[1],推测DNA甲基化、组蛋白修饰可能参与联想学习中的记忆储存[1,6];Feng等人(2016)证实,光照可通过调控组蛋白甲基化影响植物的盐胁迫转录记忆,提示光照相关的联想记忆可能存在类似机制[6]。
信号通路整合:生长素(auxin)信号系统被证实参与光信号与机械信号的整合(Halliday et al., 2009),可能是联想学习中刺激关联的核心通路[5];钙信号与活性氧(ROS)作为植物环境应答的通用信使,也被推测参与刺激信号的传导与整合[6]。
分子网络调控:无神经系统的有机体可能通过遗传调控网络的动态调整实现信息储存与处理(Bray, 2003),Sorek等人(2013)的模型预测,含双稳态元件的遗传网络可形成并储存关联记忆,为植物联想学习的分子机制提供了理论框架[6]。
4.3 研究方法的标准化探索
针对争议中暴露的实验可重复性问题,学界开始推动植物联想学习研究的方法标准化:
光照参数规范:要求详细披露所有阶段的光照条件,包括光谱组成、光照强度、光周期等(Cvrčková & Konrádová, 2022)[6],避免因光谱差异导致的结果分歧。
实验设计优化:强调采用双盲分析、扩大样本量、设置多元对照组(如Moving Light对照组),减少实验偏差(Markel, 2020)[2,3]。
多实验室联合验证:呼吁在原作者协助下,统一豌豆品种、实验装置等关键变量,开展跨实验室重复验证,消除系统性偏差[2,3,6]。
5 争议的核心分歧:三重博弈与方法论反思
5.1 核心分歧的三重维度
非条件刺激的有效性之争:Gagliano团队认为原实验中蓝光100%引发向光性,是有效US;Markel则指出1小时斜向蓝光本就难以引发强向光性,原实验结果具有特殊性;Cvrčková与Konrádová提出前期光照质量差异可能是US有效性分歧的深层原因[3,4,6]。
实验环境与材料的差异之争:包括植株间距与实验空间(全暗室vs培养箱)、豌豆品种(Massey Gem vs Green Arrow)、前期光照质量(荧光灯vs红蓝光LED)三大维度,双方对各变量的影响权重存在显著分歧[2,3,4,6]。
实验设计的严谨性之争:Markel批评原实验未采用双盲分析,对照组设计单一;Gagliano团队则认为分工记录已减少偏差,核心争议在于环境控制而非分析方法[1,2,3]。
5.2 方法论反思:植物实验的特殊性与可重复性
植物的环境敏感性:植物对光照、温度、湿度等环境变量高度敏感,即使是前期培养阶段的细微差异,也可能导致行为应答的显著不同[6],这要求植物行为研究需具备更高的变量控制精度。
可重复性的双重标准:科学研究的可重复性不仅要求实验步骤的复刻,更需要核心环境变量的一致性[3,4],植物联想学习的争议提示,未来研究需详细披露所有关键参数,包括前期培养条件。
概念定义的跨学科共识:争议也反映出“植物联想学习”的定义边界尚未明确,需结合植物的生理特性,建立区别于动物的认知行为评估标准,避免简单套用动物实验的理论框架[6,7]。
6 结论与未来展望
植物联想学习的研究历经半个多世纪的探索,从早期的初步尝试到2016年的突破性发现,再到近年来的争议与深化,已成为植物行为学与认知科学的交叉热点。Gagliano团队的原创实验首次提供了植物联想学习的系统性证据,Markel的重复实验揭示了实验条件的敏感性,Cvrčková与Konrádová的研究则拓展了光照质量历史的全新视角,三方的学术碰撞推动了研究向更严谨、更深入的方向发展。
尽管争议未平,但这些探索已打破“植物被动适应环境”的刻板印象,证明植物的行为调控机制远比想象中复杂。未来研究应聚焦三大方向:一是通过多实验室联合验证,精准复刻实验条件,明确植物联想学习的真实性;二是结合多组学技术与分子生物学方法,解析其潜在的表观遗传调控与信号通路机制;三是拓展研究对象与生态场景,验证联想学习是否为植物界的普遍现象,评估其生态适应性价值。
植物联想学习的真相尚未揭晓,但这一探索过程本身已彰显了科学研究的核心价值——通过争议与验证不断逼近真理。无论最终结论如何,该领域的研究都将重塑我们对植物认知能力与生命适应机制的理解,为生命科学的发展开辟新的视野。
重要参考文献
[1] Gagliano, M., Vyazovskiy, V. V., Borbély, A. A., Grimonprez, M., & Depczynski, M. (2016). Learning by association in plants. Scientific Reports, 6(1), 38427. https://doi.org/10.1038/srep38427
[2] Markel, K. (2020). Lack of evidence for associative learning in pea plants. eLife, 9, e57614. https://doi.org/10.7554/eLife.57614
[3] Markel, K. (2020). Response to comment on 'Lack of evidence for associative learning in pea plants'. eLife, 9, e61689. https://doi.org/10.7554/eLife.61689
[4] Gagliano, M., Vyazovskiy, V. V., Borbély, A. A., Depczynski, M., & Radford, B. (2020). Comment on 'Lack of evidence for associative learning in pea plants'. eLife, 9, e61141. https://doi.org/10.7554/eLife.61141
[5] Halliday, K. J., Martínez-García, J. F., & Josse, E.-M. (2009). Integration of light and auxin signalling. Cold Spring Harbor Perspectives in Biology, 1(6), a001586. https://doi.org/10.1101/cshperspect.a001586
[6] Cvrčková, F., & Konrádová, H. (2022). Associative learning in plants: light quality history may matter. BIOCELL, 46(3), 645-649. https://doi.org/10.32604/biocell.2022.018114
[7] Adelman, B. E. (2018). On the conditioning of plants: A review of experimental evidence. Perspectives on Behavior Science, 41(4), 431–446. https://doi.org/10.1007/s40614-018-0173-6
[8] Bray, D. (2003). Molecular networks: The top-down view. Science, 301(5641), 1864–1865. https://doi.org/10.1126/science.1089018
[9] Zhang, T., Maruhnich, S. A., & Folta, K. M. (2011). Green light induces shade avoidance symptoms. Plant Physiology, 157(4), 1528–1536. https://doi.org/10.1104/pp.111.180661
[10] Gagliano, M., Renton, M., Depczynski, M., & Mancuso, S. (2014). Experience teaches plants to learn faster and forget slower in environments where it matters. Oecologia, 175(1), 63–72. https://doi.org/10.1007/s00442-013-2873-7
[11] Sorek, M., Balaban, N. Q., & Loewenstein, Y. (2013). Stochasticity, bistability and the wisdom of crowds: A model for associative learning in genetic regulatory networks. PLOS Computational Biology, 9(9), e1003179. https://doi.org/10.1371/journal.pcbi.1003179
[12] Feng, X. J., Li, J. R., Qi, S. L., Lin, Q. F., Jin, J. B., & Hua, X. J. (2016). Light affects salt stress-induced transcriptional memory of P5CS1 in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America, 113(51), E8335–E8343. https://doi.org/10.1073/pnas.1610670114
[13] Holmes, E., & Gruenberg, G. (1965). Learning in plants. Worm Runner’s Dig, 7, 9-12.
https://wap.sciencenet.cn/blog-38998-1524316.html
上一篇:树木对星空的感知:云杉林日食生物电同步揭示的植物智慧