博文
超声驱动的铋 - 钌异质结用于肿瘤水分解气体治疗
||
超声驱动的铋 - 钌异质结用于肿瘤水分解气体治疗
摘要
癌症治疗面临着重大挑战。由于压电材料性能存在局限,压电疗法难以实现理想的肿瘤抑制效果。为此,本研究设计了两亲性聚乙二醇(PEG)修饰的Bi₂O₃/RuO₂异质结复合压电纳米球花(BROP),用于肿瘤气体治疗。多种表征方法表明,在超声(US)刺激下,BROP能高效催化水分解产生氢气(H₂)和氧气(O₂)。理论与实验研究显示,异质结构的构建提高了材料的偶极矩,显著优化了电荷分离效率与载流子迁移率,增强了压电性能,并实现了高催化活性。BROP与超声联合作用时,会损伤肿瘤细胞线粒体,诱导细胞凋亡和铁死亡,同时缓解肿瘤的缺氧微环境。在体内肿瘤模型中,该疗法实现了95.2%的肿瘤抑制率,且未对主要器官造成损伤。综上,本研究结果为压电催化气体疗法在肿瘤治疗中的应用提供了可能,并为无机金属压电声敏剂的应用奠定了基础。
本文参考文章
School of Materials and Chemistry, Institute of Bismuth Science, Shanghai Collaborative Innovation Center of Energy Therapy for Tumors, University of Shanghai for Science and Technology, Shanghai, 200093 China E-mail: yhli@usst.edu.cn
1 引言
在全球范围内,癌症治疗仍是一项艰巨的医学挑战,亟需开发更安全、更高效的治疗策略[1]。近年来,声压电疗法与治疗性压电生物材料在纳米医学领域得到了广泛研究[2]。凭借其独特的作用机制,这类材料在抗肿瘤、抗菌和免疫调节治疗领域展现出巨大的应用潜力,为生物医学治疗提供了新方向[3-6]。
肿瘤组织具有细胞增殖失控、血管生成异常和代谢废物堆积的特征,这使得肿瘤组织中的水分(H₂O)含量远高于正常组织[7,8]。这一特点为基于水分解的治疗策略提供了天然优势。压电材料(PEM)具有非中心对称晶体结构,在外加机械力(如超声)作用下可将机械能转化为化学能[3]。其产生的压电势能显著提升催化活性[9]。因此,由压电材料驱动的压电催化水分解在肿瘤治疗中具有显著优势[10,11]。分解肿瘤内的水分可降低组织间隙胶体渗透压,抑制血管内皮生长因子等促血管生成因子的分泌,阻断肿瘤细胞的糖酵解过程,增强乳酸堆积的毒性,并诱导肿瘤细胞凋亡[10,12,13]。此外,水分解产生的氢气(H₂)和氧气(O₂)也发挥着重要治疗作用:氢气具有类似化疗药物的功能,可穿透肿瘤细胞,干扰线粒体功能,切断肿瘤细胞存活与增殖所需的能量供应,加速肿瘤细胞凋亡[14-16];氧气能有效缓解肿瘤缺氧状态,改善肿瘤微环境(TME),逆转免疫抑制环境,并增强机体对肿瘤细胞的免疫活性[17,18]。因此,开发高效的压电纳米催化材料以实现肿瘤内水分子的高效分解,已成为抑制肿瘤细胞增殖的关键突破口。
然而,当前超声诱导的压电极化过程受限于压电材料载流子浓度不足和固有压电响应微弱的问题,导致电荷分离效率低下,这严重制约了压电催化效率,使得活性氧(ROS)产量较低,难以有效抑制肿瘤生长[12,19]。为提升压电性能,研究人员探索了多种策略,如构建异质结构、制造缺陷和金属离子掺杂等[3,20,21]。其中,氧空位(OVs)对压电材料的物理化学性质影响尤为显著[22]。但传统压电材料(如锆钛酸铅陶瓷)存在诸多缺陷[23]:其含有的金属元素(如Pb²⁺)可能通过体液扩散,引发细胞毒性或器官损伤[24];化学稳定性差,长期植入后易被体液侵蚀降解,并引发炎症反应[25,26];在生物组织中受超声刺激时,信号衰减快且穿透深度有限,难以用于深部肿瘤治疗[3]。尽管铋基纳米材料凭借其独特且易修饰的晶体结构、低毒性和对超声刺激的敏感性,在声压电领域展现出潜力,但仍需进一步优化才能实现高效的肿瘤治疗[27,28]。
基于上述问题,并结合对以往压电材料催化水分解研究的深入分析,本研究旨在构建一套超声催化水分解体系。我们设计了Bi₂O₃/RuO₂(BRO)压电纳米花异质结,并用两亲性聚乙二醇(PEG)对其进行包覆修饰,形成用于肿瘤治疗的复合材料(BROP)(图1)[29,30]。作为一种宽带隙半导体材料,Bi₂O₃可响应可见光,在有机污染物降解、产氢和二氧化碳还原领域具有潜在应用[31,32];而RuO₂在析氢反应(HER)和析氧反应(OER)中表现优异,且在酸性条件下具有出色的催化活性和稳定性[33]。构建Bi₂O₃/RuO₂异质结不仅能促进电荷分离,显著提升析氢和析氧活性,还能破坏晶体对称结构,形成偶极层,增强本征压电性能。在超声刺激下,异质结的内部电场与能带变化可促进电子-空穴对的解离,参与纳米颗粒表面的氧化还原反应,并驱动肿瘤组织间隙液中水分子的分解[14,17]。此外,BROP还能消耗还原型谷胱甘肽(GSH),破坏肿瘤微环境的氧化还原平衡;超声则会促进活性氧的积累和谷胱甘肽的进一步消耗,从而加剧肿瘤的氧化应激。据此,本研究旨在开发一种多功能肿瘤治疗策略,实现肿瘤内水分子的原位分解,降低组织间隙压力,进而高效抑制肿瘤生长。
![]()
图1.BROP的超声(US)触发压电催化水分解机制及其在肿瘤气体治疗中的生物学作用示意图。
2 结果与讨论
2.1 合成与表征
BROP的合成流程如图1a所示。首先通过溶剂热法合成BRO纳米颗粒,随后采用两亲性PEG对BRO纳米颗粒进行表面修饰,形成BROP纳米球花。扫描电子显微镜(SEM)和透射电子显微镜(TEM)观察显示,BRO纳米颗粒呈现规则的球花状形貌,粒径分布均匀,平均粒径约为300 nm(图1b、c)。X射线能谱(EDS)分析证实,Bi、Ru、O三种元素在BRO中均匀分布,其元素组成含量分别为44.8%、5.8%和49.4%(图1d;支持信息图S1)。电感耦合等离子体发射光谱(ICP-OES)测试表明,样品中Bi和Ru的质量分数分别为59.8%和3.2%(支持信息表S1)。此外,BRO纳米颗粒的X射线粉末衍射(XRD)图谱显示,其衍射峰与立方相Bi₂O₃(JCPDS 74-1633)和RuO₂(JCPDS 70-2662)的标准衍射峰完全匹配(图1e),证明BRO具有异质结结构。高分辨率透射电子显微镜(HRTEM)图像显示,BRO中存在多个晶格间距:经分析,平均间距为0.327 nm的晶面对应Bi₂O₃的(111)晶面,而平均间距为0.316 nm和0.255 nm的晶面则分别对应RuO₂的(110)和(101)晶面(图1f、g)。此外,HRTEM图像中的BRO还表现出晶格无序、畸变和空位特征,这些特征可能有助于提升其催化活性(支持信息图S2)[12]。
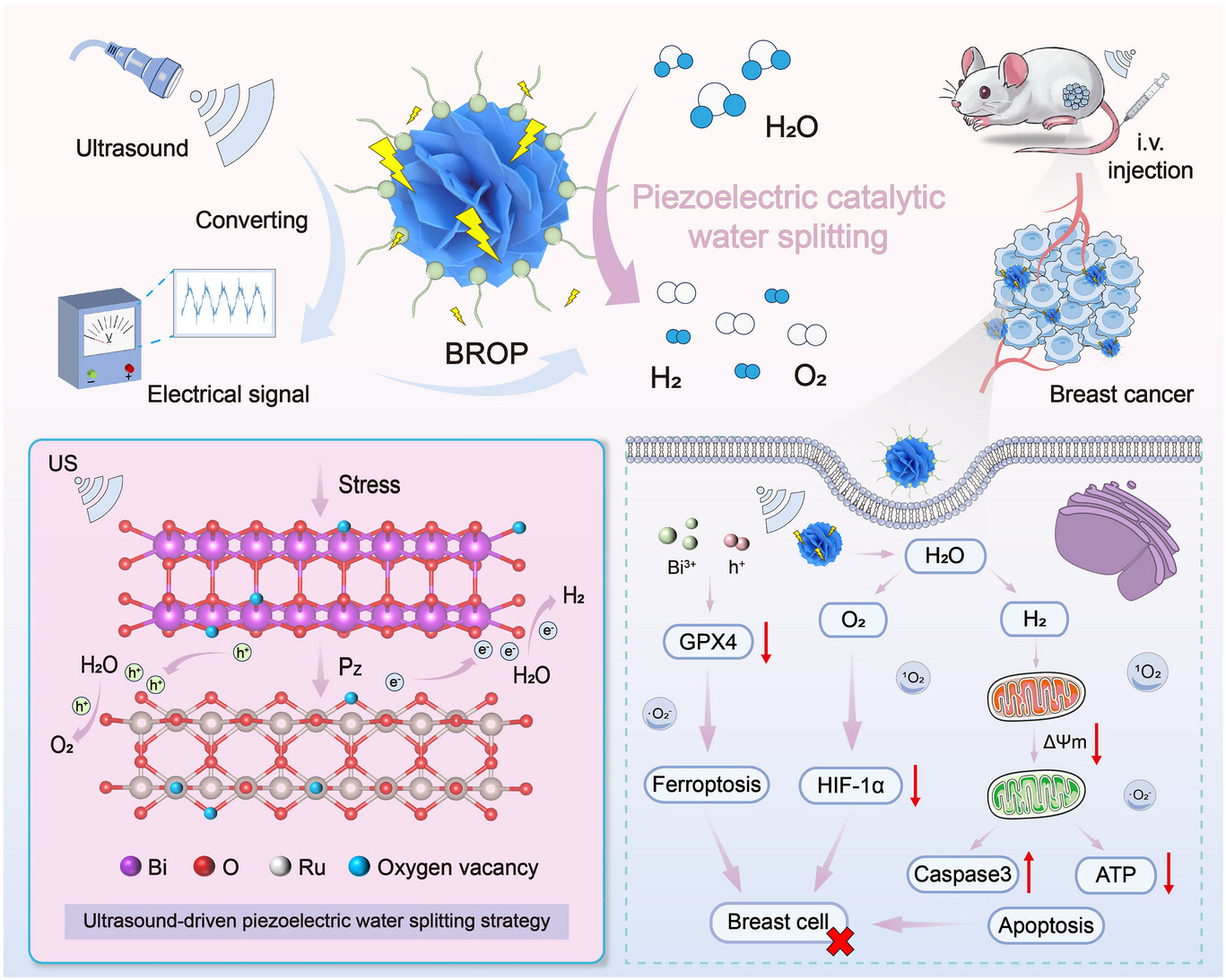
图1.a)BRO与BROP的合成示意图;b)BRO的SEM图像;c)BRO的TEM图像(插图:放大的TEM图像);d)BRO的高角环形暗场(HAADF)图像及Bi、Ru、O元素分布图谱;e)BRO的XRD图谱;f)BRO的HRTEM图像;g)BRO的晶格间距测量图;h)BRO、PEG与BROP的傅里叶变换红外(FT-IR)光谱;i)BRO与BROP的zeta电位(n=3);j)BRO与BROP的接触角测试(n=3)。数据以平均值±标准差(SD)表示。
布鲁瑙尔-埃米特-泰勒(BET)测试结果显示,Bi₂O₃的比表面积为34.298 m²/g,而异质结BRO的比表面积增至80.170 m²/g,是Bi₂O₃的2.33倍。更大的比表面积可提供更多表面活性位点,增加催化底物的吸附量和吸附概率,从而提升反应速率[34]。BRO的平均孔径为1.429 nm,大于Bi₂O₃的0.453 nm,这一特性有利于产物分子(H₂和O₂)的脱附(支持信息图S3)[35]。傅里叶变换红外(FT-IR)光谱表明,两亲性PEG已成功修饰到BRO表面(图1h;支持信息图S4),且光谱中的羟基伸缩振动峰证实BROP具有良好的亲水性[12]。Zeta电位测试显示,BRO的zeta电位为-3.36 mV,而BROP的zeta电位为-8.21 mV(图1i),进一步验证了两亲性PEG对BRO的修饰成功。接触角测试表明,BROP与水接触后呈现较小的接触角,说明其能与水充分接触,具有优异的亲水性(图1j)。动态光散射(DLS)测试显示,经两亲性PEG修饰后,BROP的水合粒径略大于BRO;此外,BROP在水和pH 7.4的磷酸盐缓冲液(PBS)中储存7天后仍保持稳定,表明其在水环境和体内生理条件下均具有良好的稳定性(支持信息图S5)。综上,多种表征方法证实,BROP是一种经两亲性PEG修饰的Bi₂O₃/RuO₂球花状异质结材料。
2.2 声催化水分解性能评估
2.2 声催化水分解性能评估
为优化BRO的催化性能,研究采用线性扫描伏安法(LSV)评估了不同Bi/Ru投料比样品的水分解活性,并拍摄了相应的透射电子显微镜(TEM)图像(支持信息图S6、S7)。在电催化过程中,析氢反应(HER)和析氧反应(OER)的过电位是衡量催化剂活性的关键指标[36]。过电位越小,意味着催化剂驱动反应所需的额外能量越低,催化活性越高。实验结果显示,当Bi/Ru投料比为2:1时,BRO在电流密度η10和η100下表现出最小的HER过电位(图2a)。在该投料比下,BRO的OER催化性能同样最优:电流密度为10 mA·cm⁻²时,过电位仅为183.36 mV;电压为1.5 V时,电流密度达到16.4 mA·cm⁻²(图2b)。通过极化曲线进一步推导塔菲尔(Tafel)图发现,Bi/Ru投料比为2:1的样品在10 mA·cm⁻²电流密度下具有更低的塔菲尔斜率。与商用RuO₂催化剂(η10=238 mV,塔菲尔斜率101.2 mV·dec⁻¹)相比(图2c)[37],该样品展现出更优异的催化动力学性能。因此,将Bi/Ru投料比调控为2:1制备的BRO,在电催化水分解反应中表现最佳。研究随后选择该投料比的样品进行后续声催化水分解性能评估。
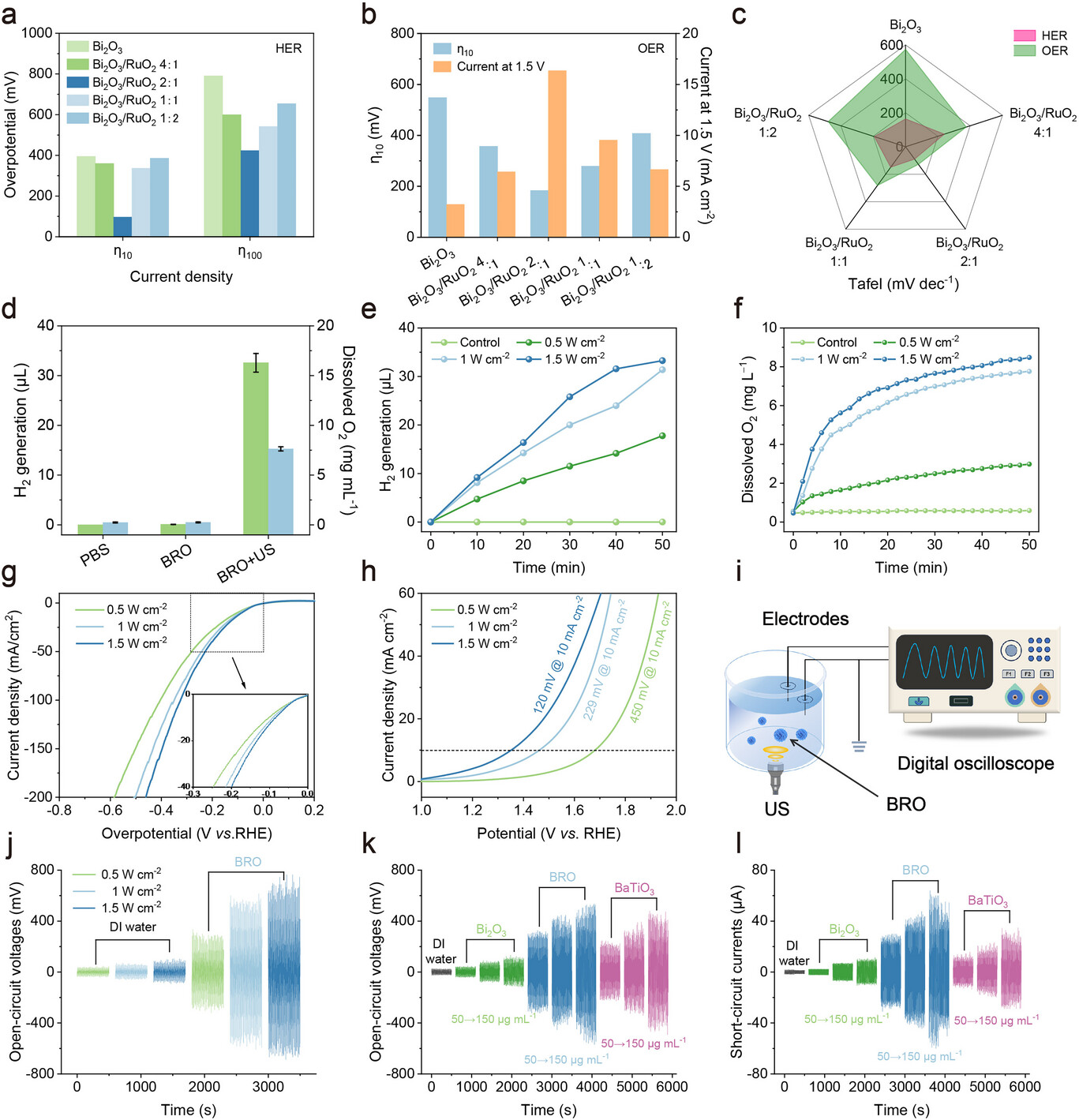
图2.a)不同投料比合成的BRO的HER活性对比;b)不同投料比合成的BRO的OER活性对比;c)基于极化曲线的塔菲尔斜率对比;d)超声(US)刺激下BRO在磷酸盐缓冲液(PBS)中的H₂和O₂产率(n=3);e)不同超声功率刺激下BRO在去离子水中的H₂产率随时间变化曲线;f)不同超声功率刺激下BRO在去离子水中的O₂产率随时间变化曲线;g)不同超声功率下BRO的HER活性对比;h)不同超声功率下BRO的OER活性对比;i)电输出信号检测装置示意图;j)不同超声功率刺激下BRO纳米球在去离子水中的开路电压波形;k)不同浓度的Bi₂O₃、BRO和钛酸钡(BaTiO₃)在去离子水中的开路电压波形;l)不同浓度的Bi₂O₃、BRO和BaTiO₃在去离子水中的短路电流波形。数据以平均值±标准差(SD)表示。
在表征超声处理下BRO的催化水分解性能时,研究对其在PBS中产生H₂和O₂的过程进行了详细评估。实验中,在1 W·cm⁻²的超声强度下对样品持续照射50分钟,分别采用气相色谱(GC)和溶解氧测定仪对生成的H₂和O₂进行定量分析(图2d;支持信息图S8)。如图2e、f所示,随着超声照射时间的延长和强度的增加,溶液中H₂生成量和溶解氧含量均呈逐渐上升趋势。这些结果直观表明,超声激发可有效促进BRO催化的水分解反应。为排除光照对水分解反应的潜在影响,研究在黑暗条件下进行了对照实验,结果显示仅在超声刺激下仍能检测到气体生成(支持信息图S8、S9),证实该体系中的水分解反应由超声激发驱动,而非光照。
为进一步验证生成气体的成分,研究利用H₂和O₂的特定化学性质开展验证实验。H₂与部分益生菌类似,可作为抗氧化生物分子清除羟基自由基(•OH)[38,39]。亚甲基蓝(MB)是检测H₂的快速探针,在铂(Pt)纳米颗粒催化下可被H₂还原为无色产物[40]。为测定H₂产率,研究向溶液中加入适量还原型谷胱甘肽(GSH)以模拟肿瘤微环境(TME),同时在测试体系中引入甲醇作为猝灭剂以消除•OH的干扰。对比单纯超声处理组与BRO+超声处理组发现,MB探针组在超声照射下664 nm处的吸光度值逐渐下降,表明BRO+超声照射组能够产生H₂,同时也说明作为空穴牺牲剂的GSH有助于H₂生成(支持信息图S10)。随后,研究进一步验证了不同超声功率下BRO溶液的H₂生成情况。如支持信息图S11所示,BRO的H₂生成效率与超声功率和时间呈显著线性相关。综上,可确认该体系中确实生成了H₂。研究还采用二氯三(4,7-二苯基-1,10-菲咯啉)钌(II)(Ru(bpy)₃Cl₂)作为氧敏感探针,其在605 nm处的荧光强度随O₂生成量的增加而降低。超声激发下BRO纳米颗粒的荧光信号显著减弱(支持信息图S12),有力证实了O₂的生成。
为深入研究超声功率对水分解反应的影响,研究通过电化学测试进一步验证。当超声功率从0.5 W·cm⁻²逐渐增加至1.5 W·cm⁻²时,BRO在相同电流密度下的过电位呈逐渐下降趋势(图2g)。该现象表明,超声处理可有效驱动水分解反应,且更高的超声功率能显著提升BRO的HER活性。类似地,图2h所示的实验结果呈现相同趋势,证实BRO的OER动力学过程显著加快,催化性能得到提升。在超声处理的作用下,BRO的OER过程可能激活了吸附演化机制(AEM)(支持信息图S13)。值得注意的是,在1.5 W·cm⁻²的超声功率下,BRO的OER过电位显著降至120 mV(电流密度10 mA·cm⁻²),这进一步凸显了高超声功率下BRO优异的催化动力学特性。
明确BRO在声催化水分解过程中的能量转换机制也至关重要。研究采用数字示波器研究其在超声条件下的声电响应特性(图2i)。实验以1.0 MHz的超声激发BRO,结果显示:随着超声功率的增加,BRO的开路电压呈上升趋势,且与超声强度呈正相关(图2j)。此外,在相同激发功率下,随着BRO纳米颗粒浓度的增加,开路电压从352 mV逐渐升至654 mV并趋于稳定,其输出电压显著高于常见压电材料(PEM)钛酸钡(BaTiO₃)(图2k)。同样,短路电流的变化趋势与开路电压一致:当BRO浓度为100 μg·mL⁻¹时,可产生约40 μA的电流(图2l)。这些声电性能表征结果证实,BRO在超声刺激下会产生压电势,为声催化水分解过程中机械能-电能-化学能的高效转换提供了直接实验证据。
2.3 压电性能评估与机制分析
以往研究表明,超声处理产生的机械能可通过压电效应克服水分解过程中的固有动力学势垒,从而促进HER和OER的发生[11,12]。在压电材料研究中,非中心对称结构的增强能有效产生极化电荷,促进载流子分离,进而触发声电效应[41]。因此,压电性能和半导体特性是评估材料声催化活性的关键参数。本研究首先验证了BRO在超声刺激下会周期性产生压电感应电势(图3a),随后明确了BRO声催化水分解的机制。
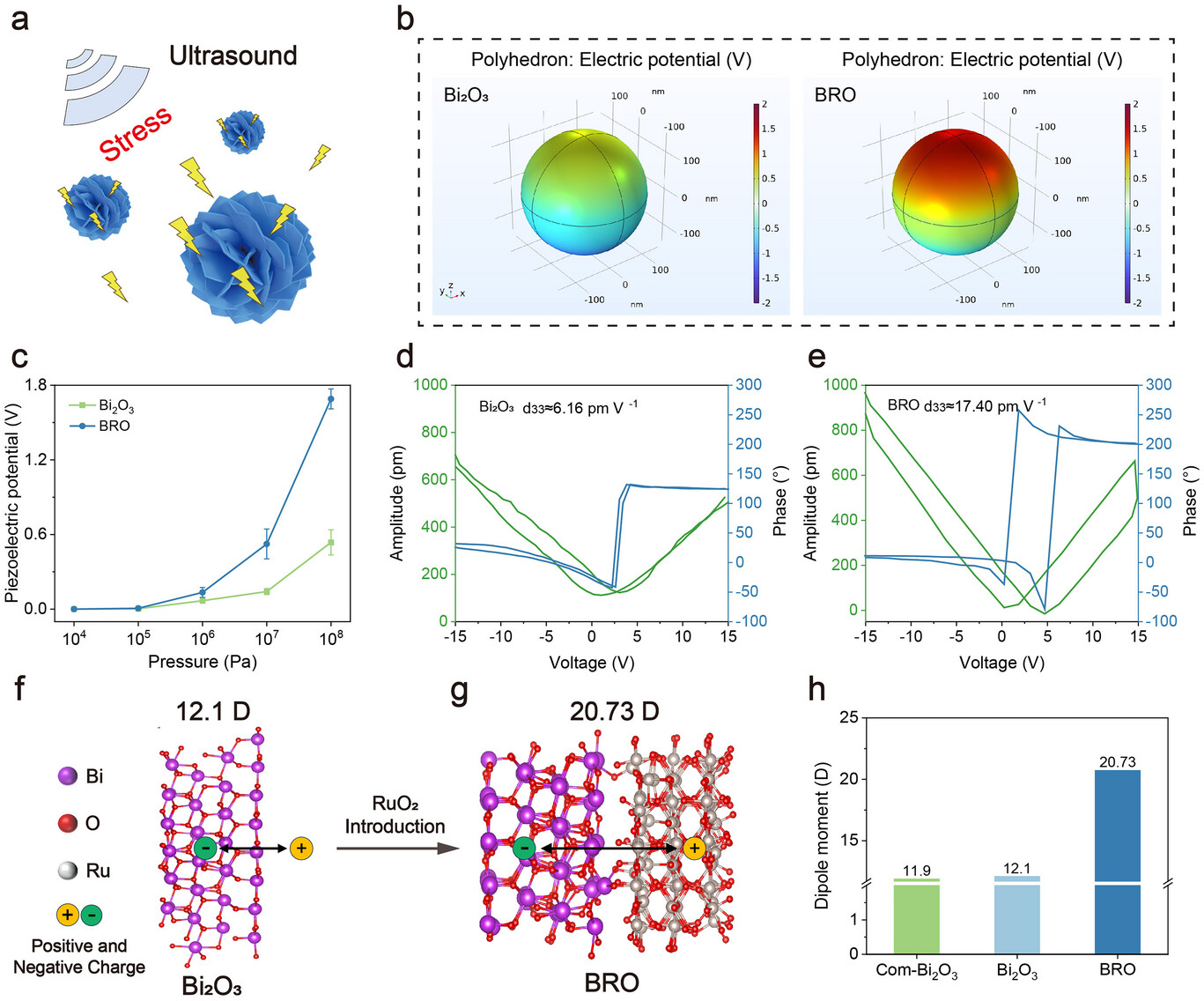
图3.a)超声触发BRO纳米球产生表面压电势的示意图;b)所制备样品电势分布的COMSOL模拟图;c)压力在10⁴~10⁸ Pa范围内时Bi₂O₃和BRO的表面压电势(n=3);d)Bi₂O₃的振幅-电压蝴蝶曲线及相应相位滞后回线;e)BRO的振幅-电压蝴蝶曲线及相应相位滞后回线;f)Bi₂O₃的模拟电荷分离示意图;g)BRO的模拟电荷分离示意图;h)商用Bi₂O₃、溶剂热合成Bi₂O₃与BRO的偶极矩对比。数据以平均值±标准差(SD)表示。
从宏观力学和电学响应角度,研究采用COMSOL Multiphysics软件进行有限元分析(图3b),以确定不同压力强度下Bi₂O₃与BRO的感应电势差异。模拟结果显示,与Bi₂O₃相比,BRO在不同强度的超声压力下均能产生更强的感应电势,且感应电势与压力呈指数关系(图3c)。该现象表明,在超声作用下,BRO内部产生更大的应变,导致电荷分布发生更显著变化,从而产生更高的表面压电势,体现出BRO优异的宏观压电性能。
为分析宏观性能差异的内在原因,研究采用压电响应力显微镜(PFM)精确测量Bi₂O₃和BRO的压电性能。结果显示,当施加-15 V和15 V的偏压时,两者均呈现出明显的蝴蝶形振幅曲线和约180°的相位曲线,表明外电场会引起材料发生应变,并表现出可逆极化特性(图3d、e)。通过振幅衰减回线进一步计算发现,BRO的压电系数d33达到17.40 pm·V⁻¹,约为Bi₂O₃(6.16 pm·V⁻¹)的2.8倍。已知Bi₂O₃本身具有萤石型立方对称晶体结构,不利于压电性能的产生[44];但氧空位(OVs)的存在会诱导结构变化并降低对称性,从而使材料产生一定的压电性能[22]。结合Bi₂O₃和BRO的电子顺磁共振(EPR)分析,证实了强氧空位信号(g=2.0026)的存在(支持信息图S14)。BRO中异质结的构建进一步扰乱了结构对称中心,从宏观结构层面显著提升了材料的压电性能。
在微观电子结构层面,研究采用密度泛函理论(DFT)计算Bi₂O₃和BRO的偶极矩变化(图3f、g),以明确不对称性诱导极性变化及压电增强的机制[42]。偶极矩的变化反映了材料利用偶极场驱动极化、促进电荷分离的潜在能力。结果显示,Bi₂O₃的偶极矩受氧空位(OVs)导致的正负电荷中心不重合影响,初始值为12.1 D,高于商用Bi₂O₃的11.9 D;当引入RuO₂形成异质结后,BRO的偶极矩显著提升至20.73 D(图3h)。这种提升源于微观层面多因素的协同作用:Ru的d轨道离域特性,结合Bi的6s²孤对电子易极化的特点,形成了强界面偶极[43,44];RuO₂具有类近半金属特性[45],可加速电荷转移[46],促使Bi₂O₃/RuO₂异质结两侧发生能带弯曲,形成内建电场(IEF),进一步增强BRO的本征极化,最终导致偶极矩显著增加。
为进一步证实偶极矩变化与压电性能提升的关联性,研究通过恒电流充放电测试对比Bi₂O₃和BRO的面电容值与相对介电常数,以量化BRO的电荷存储能力。结果显示,与Bi₂O₃相比,BRO具有更多氧化还原反应活性位点、更高效的电荷存储机制,且在电场下更易极化(支持信息图S15)。BRO更高的介电常数使其能形成更多偶极子,具备更强的电能存储能力[47]。当超声压力作用于BRO表面时,产生的应变会引起材料内部电荷分布变化,诱导极化响应。作为衡量分子极性的重要参数,偶极矩的增加直接反映了材料极化程度的增强,进而提升压电性能[48,49]。
2.4 半导体性能与机制分析
随后,研究通过分析BRO的半导体特性,进一步探究声催化水分解机制。BRO在不同扫描速率下的循环伏安(CV)曲线呈现典型的矩形特征,体现出理想的电容行为。在超声激活下,BRO的电流密度显著增加,对应双电层电容(Cdl)增强,且活性位点暴露程度提高(图4a;支持信息图S16),表明BRO的声催化性能可促进电荷在材料表面积累,提升材料与溶液间的电荷交换效率,从而优化电荷转移过程。
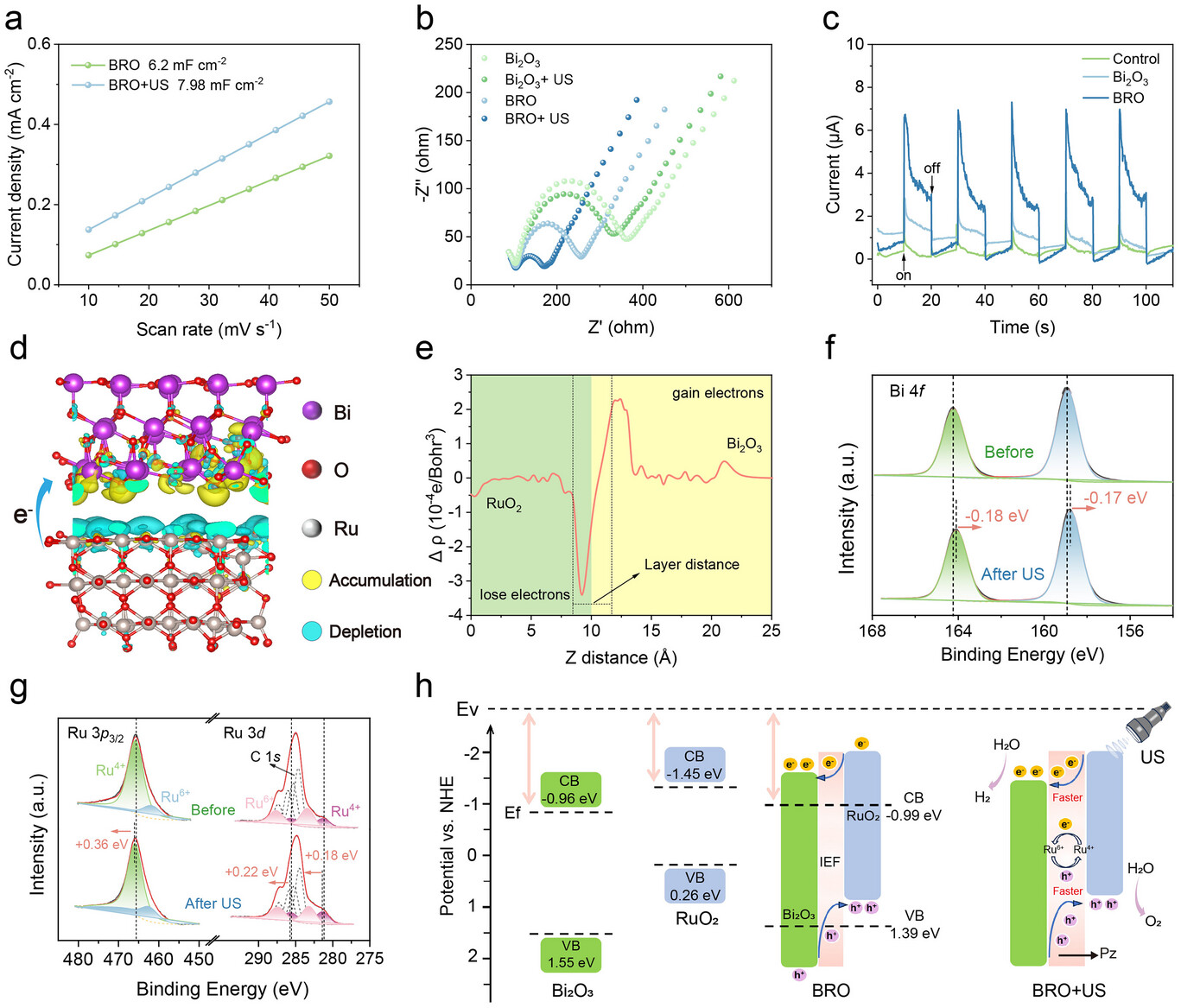
图4.a)超声处理与未处理的BRO双电层电容对比;b)Bi₂O₃和BRO在超声处理前后的电化学阻抗谱(EIS)图;c)Bi₂O₃和BRO在5个“开/关”循环中的声电电流响应;d)模拟BRO中电荷重排的示意图(黄色代表电荷聚集,蓝色代表电荷缺失);e)一维差分电荷密度图;f)超声处理前后BRO的Bi 4f X射线光电子能谱(XPS);g)超声处理前后BRO的Ru 3p和Ru 3d XPS谱;h)Bi₂O₃、RuO₂和BRO的能带图及超声压电催化机制示意图。
为进一步表征BRO的电荷转移机制,研究采用电化学阻抗谱(EIS)和瞬态声电电流谱(i-t曲线)开展研究。作为声催化的核心过程,压电材料(PEM)中电子-空穴对的产生、分离及迁移效率直接决定其催化活性[9]。EIS测试显示,BRO样品的阻抗弧半径显著小于Bi₂O₃,表明其电荷转移电阻更低;经超声激活后,BRO的电荷转移能力进一步增强(图4b)。此外,i-t曲线测试显示,超声作用下BRO的瞬态声电电流显著高于Bi₂O₃,证实超声处理可有效提高载流子生成量(图4c)。同时,计时电流法测试显示,BRO在100 mV和200 mV电压下运行15000 s以上后,活性衰减较小(支持信息图S17),体现出良好的稳定性。这些结果共同表明,超声激活下BRO的电荷转移能力和载流子动力学特性显著优于Bi₂O₃。
为揭示上述性能差异的微观本质,研究结合DFT模拟与X射线光电子能谱(XPS)分析,明确BRO的电子结构变化。DFT模拟显示,Bi₂O₃与RuO₂接触后,界面处发生电荷重排,形成从RuO₂指向Bi₂O₃的内建电场(IEF)(图4d、e)。XPS表征进一步验证了这一结论:当Bi₂O₃与RuO₂形成BRO异质结时,Bi 4f轨道特征峰发生0.1 eV的负向偏移,表明Bi与Ru原子间存在电子相互作用;经超声催化水分解反应后,Bi 4f₅/₂和Bi 4f₇/₂峰分别负向偏移0.18 eV和0.17 eV,Bi的电子云密度显著增加;与之相反,Ru的+6价态比例升高,Ru 3p₃/₂和Ru 3d轨道特征峰发生正向偏移,电子密度降低(图4f、g;支持信息图S18)。综上,结果表明在催化过程中,电子从Ru中心转移至Bi中心,进一步增强内建电场。
基于上述电子结构分析,研究通过固体漫反射光谱和价带光谱确定Bi₂O₃、RuO₂和BRO的能带结构(支持信息图S19)。由于Bi₂O₃与RuO₂的电子亲和能和电离能存在差异,界面两侧材料的费米能级不同,从而形成稳定的内建电场[50]。在超声刺激下,BRO产生的压电势引发能带弯曲,内建电场为电子-空穴对的定向分离提供驱动力:电子沿内建电场方向在Bi₂O₃中积累,催化H₂O还原生成H₂;空穴迁移至RuO₂,氧化H₂O并析出O₂(图4h)。此外,BRO的电势满足生成单线态氧(¹O₂)和超氧阴离子(•O₂⁻)的条件(相对于标准氢电极(NHE),O₂/•O₂⁻的电势为-0.33 eV)[35]。研究采用¹O₂探针9,10-二苯基蒽(DPA)和•O₂⁻探针氯化硝基四氮唑蓝(NBT)验证活性氧(ROS)的生成(支持信息图S20)。上述结果共同进一步证实,BRO在声催化水分解过程中具备高效的电荷分离与催化转化能力。
2.5 细胞水平治疗效果评估
研究通过系统实验探究BROP在肿瘤治疗领域的应用潜力。首先,生物透射电子显微镜(Bio-TEM)成像显示,BROP可有效分布于4T1小鼠乳腺癌细胞的细胞质和囊泡中(图5a)。BROP-RhB体系采用罗丹明(RhB)荧光标记,进一步证实4T1细胞与BROP共孵育12 h后,对BROP的摄取达到饱和状态(支持信息图S21)。
研究采用细胞计数试剂盒8(CCK-8)评估BROP的生物相容性:将不同浓度的BROP与4T1肿瘤细胞、NIH-3T3小鼠胚胎成纤维细胞共孵育48 h后,结果显示NIH-3T3细胞活力未受显著抑制,充分证实BROP的生物安全性;当BROP浓度超过80 μg·mL⁻¹时,4T1细胞活力显著低于NIH-3T3细胞,推测其对肿瘤细胞的选择性毒性可能与消耗细胞内还原型谷胱甘肽(GSH)密切相关(图5b)。此外,引入超声照射后,细胞杀伤效果显著增强:对比不同超声功率下的半数抑制浓度(IC₅₀)发现,与0.5 W·cm⁻²相比,1 W·cm⁻²和1.5 W·cm⁻²功率下的IC₅₀分别降低55.3%和60.2%(图5c、d),表明超声与BROP的协同作用可有效提升肿瘤细胞杀伤效率。

图5.a)材料内化12 h后4T1细胞的TEM图像(黄色箭头指示线粒体,橙色箭头指示BROP);b)NIH-3T3细胞和4T1细胞与BROP共孵育48 h后的细胞活力(n=5);c)不同超声功率处理后4T1细胞的活力;d)不同超声功率下的IC₅₀值(n=5);e)不同处理后,经亚甲基蓝(MB)染色的4T1细胞内H₂生成定性分析;f)采用缺氧荧光探针检测4T1细胞内O₂生成的共聚焦图像;g)JC-1染色结果;h)JC-1线粒体膜电位流式细胞术分析;i)不同处理后4T1细胞线粒体的代表性TEM图像(黄色箭头指示健康线粒体,红色箭头指示受损线粒体);j)不同处理后4T1细胞中凋亡相关蛋白的蛋白质印迹(Western blot)分析;k)不同处理后4T1细胞中缺氧相关蛋白的Western blot分析。数据以平均值±标准差(SD)表示。采用单因素方差分析(ANOVA)结合Tukey多重比较事后检验确定统计学显著性,其中*p<0.05,**p<0.01,***p<0.001,无显著性差异(ns)。
为明确BROP联合超声杀伤肿瘤细胞的内在机制,研究对细胞内关键活性物质的生成进行表征。分别采用亚甲基蓝(MB)探针和Ru(bpy)₃Cl₂探针监测细胞内H₂和O₂的生成[17,40]。结果显示,经BROP+超声处理的4T1细胞中,MB探针的蓝色显著褪色,表明BROP在细胞内高效生成H₂(图5e);同时,该组细胞中Ru(bpy)₃Cl₂探针的红色荧光强度显著降低,证实O₂的原位生成,表明超声激发下BROP可有效缓解肿瘤细胞的缺氧微环境(图5f)。
由于H₂对线粒体具有特异性靶向能力[51],研究采用线粒体膜电位检测试剂盒(JC-1)检测线粒体膜电位(ΔΨₘ):正常情况下,线粒体因膜电位较高,会形成JC-1红色荧光聚集体;当线粒体膜电位降低时,JC-1以绿色荧光单体形式存在,提示线粒体功能异常。结果显示,单独BROP组或单独超声组仅导致线粒体绿色荧光轻微增加,而BROP+超声联合处理组则出现显著的绿色荧光信号,反映膜电位显著下降(图5g;支持信息图S22)。通过流式细胞术对细胞去极化比例进行定量分析,进一步验证了上述趋势:BROP+超声组4T1细胞的去极化比例显著升高,同时细胞内三磷酸腺苷(ATP)水平降低(图5h;支持信息图S23),表明BROP生成的H₂有效损伤4T1细胞的线粒体。此外,TEM观察显示,经BROP+超声处理的肿瘤细胞线粒体严重受损,呈现出线粒体收缩、破裂、嵴消失等特征性形态变化(图5i)。
研究采用Western blot检测4T1细胞中凋亡相关和缺氧相关关键蛋白(包括前胱天蛋白酶3(pro-Caspase 3)和缺氧诱导因子1α(HIF-1α))的表达水平。结果显示,BROP+超声处理显著下调4T1细胞中pro-Caspase 3和HIF-1α蛋白的表达水平,证实超声刺激下的BROP通过诱导肿瘤细胞凋亡、缓解肿瘤缺氧微环境等多种途径,实现对肿瘤细胞的高效杀伤(图5j、k;支持信息图S24)。
2.6 细胞死亡机制分析
在深入研究BROP联合超声诱导4T1细胞死亡的机制过程中,研究重点关注细胞内活性氧(ROS)水平调控、还原型谷胱甘肽(GSH)代谢与铁死亡通路激活之间的关联。由于线粒体损伤和O₂生成均能通过不同途径影响细胞内ROS稳态[35,40,52],研究选用2',7'-二氯二氢荧光素二乙酸酯(DCFH-DA)作为探针,监测细胞内ROS水平变化。结果显示,经BROP+超声处理的4T1细胞中ROS显著生成(图6a、b),表明超声刺激下的BROP促使细胞内ROS水平超过毒性阈值,最终通过加剧氧化应激诱导细胞凋亡。
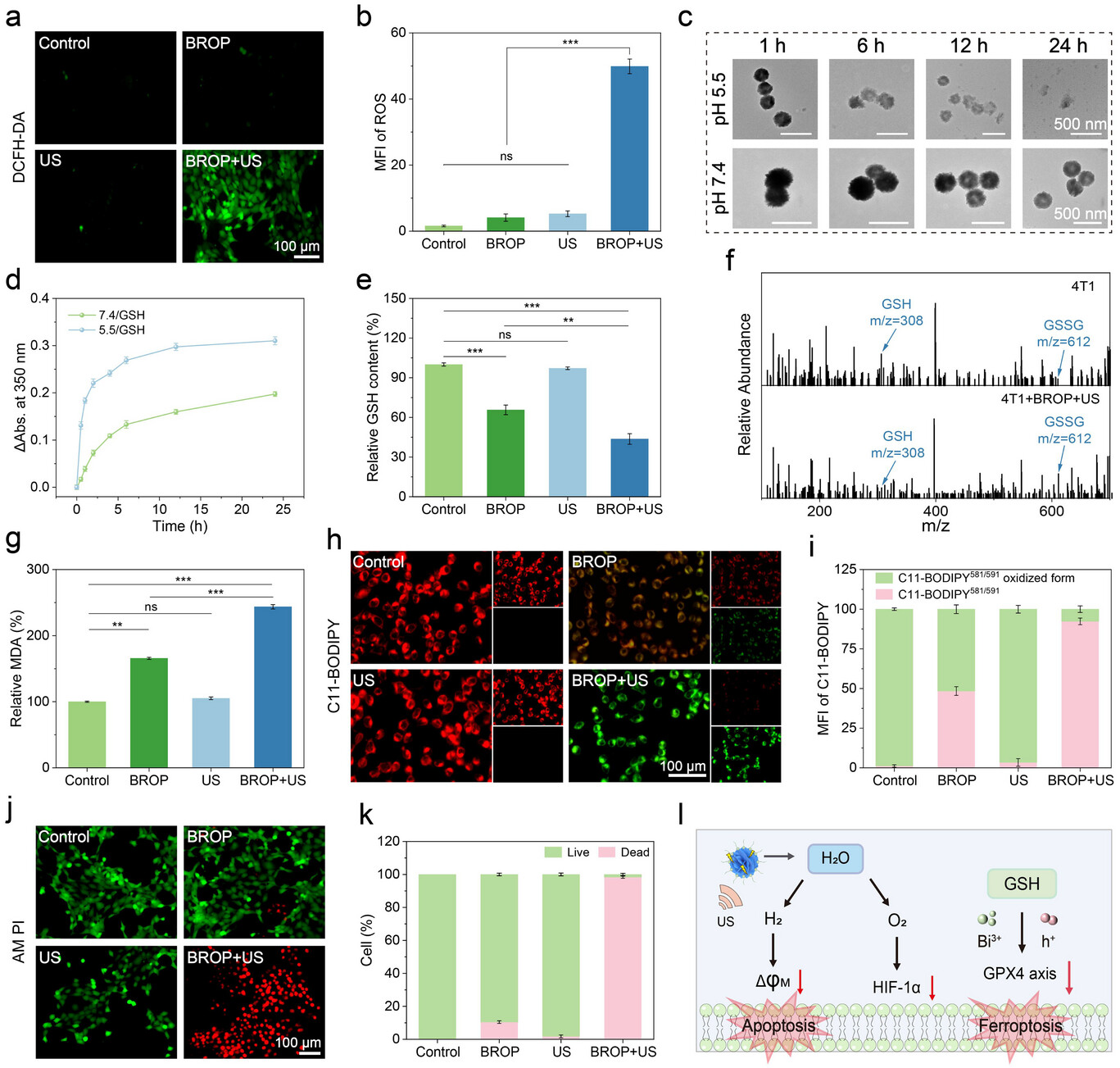
图6.a)经2',7'-二氯二氢荧光素二乙酸酯(DCFH-DA)染色的4T1细胞荧光图像;b)活性氧(ROS)荧光强度分析;c)BROP与谷胱甘肽(GSH)溶液在pH 5.5或pH 7.4条件下、不同时间点共孵育的透射电子显微镜(TEM)图像;d)相应的随时间变化的吸收光谱;e)不同处理后4T1细胞内的相对GSH含量(n=5);f)BROP+超声(US)处理后4T1细胞裂解液的高效液相色谱(HPLC)-质谱图;g)不同处理后4T1细胞内的相对丙二醛(MDA)含量(n=5);h)各组经C11-BODIPY 581/591染色的4T1细胞图像;i)荧光定量分析(n=3);j)不同处理下4T1细胞的钙黄绿素-乙酰氧基甲酯/碘化丙啶(Calcein-AM/PI)染色结果;k)荧光定量分析(n=3);l)超声刺激下BROP诱导4T1细胞凋亡、缓解缺氧及发生铁死亡的机制示意图。数据以平均值±标准差(SD)表示。采用单因素方差分析(ANOVA)结合Tukey多重比较事后检验确定统计学显著性,其中*p<0.05,**p<0.01,***p<0.001,无显著性差异(ns)。
进一步研究发现,肿瘤微环境(TME)中高表达的GSH可通过维持谷胱甘肽过氧化物酶4(GPX4)的活性来抑制铁死亡过程[53-55]。为明确BROP消耗GSH的机制,研究将BROP置于不同生理条件的环境中孵育,并通过TEM和吸光度法进行分析。结果显示,在含GSH的pH 5.5磷酸盐缓冲液(PBS)中,BROP发生快速降解,且其在350 nm处的吸光度显著升高(图6c、d;支持信息图S25)。该现象证实,酸性条件可加速Bi³⁺与GSH巯基之间的配位反应,形成Bi(GS)₃复合物,这一过程既能有效消耗GSH,也能触发铋基纳米材料的生物降解[35]。在超声刺激下,BROP产生的空穴(h⁺)与Bi³⁺协同作用,进一步增强对GSH的消耗能力[56]。采用5,5'-二硫代双(2-硝基苯甲酸)(DTNB)进行定量测定显示,与对照组相比,BROP+超声处理组细胞内GSH含量降低了56%(图6e)。高效液相色谱-质谱(HPLC-质谱)分析结果也显示,BROP与4T1细胞共孵育48 h后,细胞内GSH与氧化型谷胱甘肽(GSSG)的比例显著降低(图6f),进一步表明细胞内氧化还原平衡被破坏[57]。此外,脂质过氧化(LPO)是铁死亡的关键标志物[58],其产物MDA在BROP+超声处理组中的含量显著高于其他组(图6g)。细胞内Fe²⁺水平升高是铁死亡的重要特征之一,BROP+超声处理组中Fe²⁺水平显著升高,表明治疗过程中出现铁过载,进而触发铁依赖性铁死亡(支持信息图S26)。C11-BODIPY(脂质过氧化传感器)染色结果进一步证实了这一结论(图6h、i),表明铁死亡通路被成功激活。
为直观展示BROP对肿瘤细胞存活状态的影响,研究采用Calcein-AM/PI染色,结果显示BROP+超声处理组中红色荧光标记的死亡细胞比例显著高于其他组(图6j、k;支持信息图S27)。这些结果表明,在超声刺激下,BROP通过压电催化水分解产生H₂和O₂,既诱导细胞凋亡、缓解肿瘤缺氧微环境,又通过大量生成ROS破坏细胞氧化还原平衡,同时Bi³⁺与超声空化协同消耗GSH,最终触发铁死亡级联反应(图6l)。这种多机制协同作用模式,为BROP应用于肿瘤治疗提供了更全面的理论依据。
2.7 肿瘤模型中的疗效评估
为验证BROP在体内的肿瘤治疗潜力,研究对其体内分布特征、治疗效果及安全性进行了系统研究。为准确定位BROP在体内的最佳肿瘤蓄积时间和治疗窗口,研究将近红外荧光染料IR780负载到BROP上,制备BROP-IR780荧光纳米复合物。通过尾静脉将BROP-IR780注射到BALB/c小鼠体内,利用体内荧光成像技术动态监测其在肿瘤和主要器官中的时空分布。根据肿瘤部位荧光信号的动态变化发现,注射后12 h是BROP在肿瘤组织中富集的峰值时间,因此选择该时间点作为超声治疗的最佳时机。生物分布分析显示,肝脏和肾脏中的荧光强度在注射后12 h达到峰值,随后逐渐降低,表明BROP主要通过肝脏和肾脏代谢排出(支持信息图S28)。这些结果为后续治疗方案的设计提供了依据。
基于上述分布特征,研究设计了体内治疗实验以评估BROP的肿瘤抑制能力(图7a)。将BALB/c小鼠随机分为4组(n=4),构建皮下4T1肿瘤模型,各组分别为对照组(注射PBS)、BROP组、超声组和BROP+超声组。采用光声成像(PAI)检测肿瘤内超声催化产生的气体。根据支持信息图S29所示的氧合血红蛋白图像,对照组肿瘤的氧合血红蛋白信号较弱,而静脉注射BROP的小鼠在1 W·cm⁻²超声照射10 min后,肿瘤内血氧水平显著升高。在14天治疗周期内,结果显示各组小鼠体重无显著差异,仅BROP+超声组体重增长相对缓慢(图7b)。研究推测体重增长差异是由肿瘤抑制作用导致的。对肿瘤体积的进一步分析显示,对照组、BROP组和超声组的肿瘤呈线性生长,而BROP+超声组的肿瘤生长受到显著抑制(图7c、d)。治疗结束后,对肿瘤组织进行称重和拍照,结果显示对照组肿瘤快速生长,BROP组和超声组肿瘤体积仅轻微缩小,而BROP+超声组肿瘤体积和质量显著降低,肿瘤抑制率高达95.2%(图7e、f)。
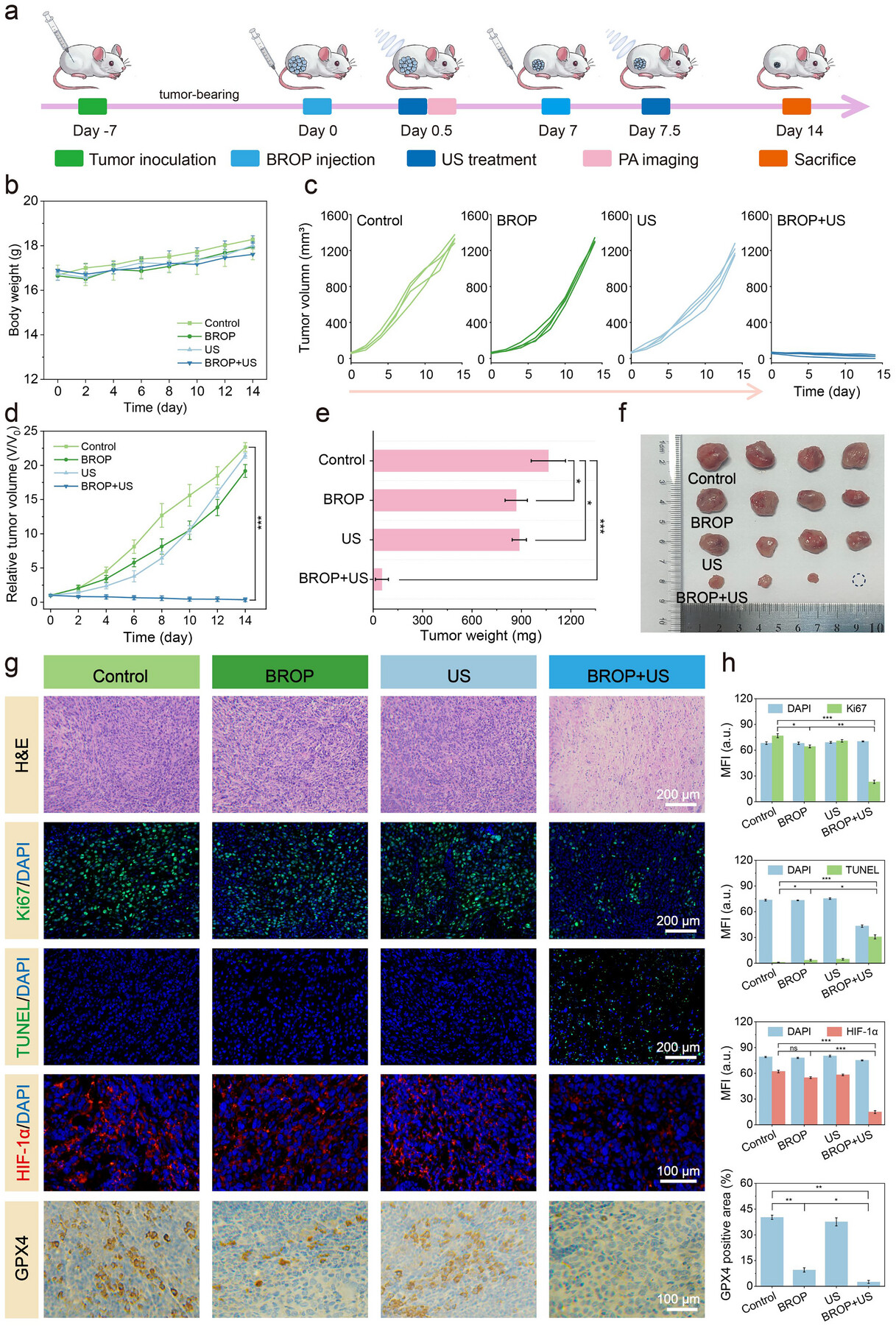
图7.a)体内肿瘤治疗方案;b)小鼠体重变化曲线(n=4);c)治疗期间各组荷瘤小鼠的肿瘤生长曲线;d)治疗期间各组荷瘤小鼠的肿瘤相对生长曲线(n=4);e)治疗14天后各组的平均肿瘤重量(n=4);f)各组肿瘤的数码照片(圆形虚线标注经相应治疗后治愈的肿瘤);g)各组肿瘤的苏木精-伊红(H&E)、Ki-67、末端脱氧核苷酸转移酶介导的dUTP缺口末端标记(TUNEL)、缺氧诱导因子1α(HIF-1α)及GPX4染色切片;h)图7g的相应定量分析(n=3)。数据以平均值±标准差(SD)表示。采用单因素方差分析(ANOVA)结合Tukey多重比较事后检验确定统计学显著性,其中*p<0.05,**p<0.01,***p<0.001,无显著性差异(ns)。
H&E染色结果显示,BROP+超声组肿瘤组织出现显著形态学改变;Ki-67染色表明该组肿瘤细胞增殖受到显著抑制。TUNEL、HIF-1α和GPX4染色进一步证实,在超声照射下,BROP诱导肿瘤细胞发生广泛损伤和凋亡,从而显著改善肿瘤组织的氧合状态,逆转缺氧微环境,并促进肿瘤细胞发生铁死亡。这些发现与整体治疗效果高度一致(图7g、h)。对主要器官组织的H&E染色显示,各器官均无明显形态学变化(支持信息图S30)。结合血液学和组织学分析结果(支持信息图S31–S33),这些结果表明BROP通过压电催化实现了高效肿瘤抑制,且对肾脏、肝脏等代谢器官无影响,具有良好的体内长期安全性。因此,体内研究证实,在超声刺激下,BROP通过压电催化水分解这一独特机制实现了高效肿瘤抑制,同时未表现出毒性作用。
要不要我帮你整理一份**文中体内实验核心数据(如肿瘤抑制率、器官安全性指标等)的检测方法、结果及临床意义对应表**?这样后续总结BROP的体内治疗价值时,能更清晰地关联实验证据与应用潜力。
3 结论
本研究制备了两亲性聚乙二醇(PEG)包覆的Bi₂O₃/RuO₂异质结纳米球花(BROP),该材料在超声(US)处理下展现出独特的压电响应特性,可用于水分解。通过数字示波器、线性扫描伏安法(LSV)等技术证实,BROP在超声刺激下不仅具有电输出能力,还能高效催化肿瘤组织中的水分解,实现气体治疗。
结合实验表征与密度泛函理论(DFT)计算,本研究揭示了异质结构构建提升BROP性能的核心机制:异质结的形成显著提高了材料的偶极矩,优化了电荷分离效率与载流子迁移能力,从而大幅增强其压电催化活性。
在肿瘤治疗应用中,BROP表现出优异疗效。超声激活下,BROP产生的压电效应可有效电解肿瘤组织间隙液中的水,持续生成氢气(H₂)和氧气(O₂)。这一过程不仅能通过直接损伤肿瘤细胞线粒体诱导细胞凋亡,还能显著缓解肿瘤微环境(TME)的缺氧状态,并高效消耗高表达的还原型谷胱甘肽(GSH)以诱导铁死亡,通过多机制协同抑制肿瘤生长。
体内实验结果显示,BROP联合超声治疗的肿瘤抑制率高达95.2%,且未对主要器官造成明显损伤,体现出良好的生物安全性。本研究为无机金属压电声敏剂在超声驱动水分解中的应用开辟了新方向,也为新型肿瘤压电催化治疗提供了理论依据。
4 实验部分
4.1 Bi₂O₃/RuO₂(BRO)的制备
1. 将硝酸铋五水合物(Bi(NO₃)₃·5H₂O,48.5 mg,0.1 mmol)溶解于20 mL乙二醇溶液中;
2. 另取三氯化钌(RuCl₃,10.4 mg,0.05 mmol)溶解于2 mL去离子水中;
3. 将三氯化钌溶液逐滴加入硝酸铋五水合物溶液中,以1000转/分钟(rpm)的转速搅拌30分钟;
4. 加入100 mg聚乙烯吡咯烷酮(PVP),在相同搅拌条件下继续反应2小时;
5. 将混合溶液转移至三颈烧瓶中,向体系内通入氮气(N₂),并在氮气氛围下将溶液加热至160℃,保温反应3小时后停止加热与搅拌;
6. 待溶液冷却至室温后收集产物,以10000 rpm的转速离心10分钟,所得沉淀用去离子水与乙醇交替洗涤3次;
7. 干燥后得到目标产物,同时制备出不同投料比的Bi₂O₃/RuO₂复合纳米颗粒及纯Bi₂O₃纳米颗粒。
4.2 Bi₂O₃/RuO₂-PEG(BROP)的制备
将5 mg BRO与20 mg两亲性PEG溶解于10 mL去离子水中,以600 rpm的转速搅拌12小时,随后通过蒸发去除溶剂,得到BROP。
4.3 超声激活下BRO电输出性能的测定
1. 配制不同浓度的BRO水溶液(20 mL);
2. 将超声探头(面积4 cm²)置于烧杯下方以产生超声波,超声治疗仪器(WED-100,中国)的输出参数设置为:频率1.0 MHz,占空比50%;
3. 测定BRO电输出信号时,将正负电极浸入溶液中,电极另一端连接至数字示波器(UPO2202,中国)。
4.4 理论计算
偶极矩与差分电荷的计算基于DFT框架,采用投影缀加平面波(PAW)方法,通过Vienna从头算模拟软件包(VASP)实现。交换关联势选用Perdew-Burke-Ernzerhof(PBE)广义梯度近似(GGA),平面波截断能设为450 eV。迭代求解Kohn-Sham方程时,能量收敛标准设为10⁻⁵ eV,布里渊区积分采用2×2×1 k点网格。所有结构均进行弛豫优化,直至原子残余力小于0.02 eV·Å⁻¹。
4.5 细胞培养与分组
乳腺癌4T1细胞与小鼠胚胎成纤维细胞NIH-3T3均购自中国科学院上海生命科学研究院。细胞培养于含10%胎牛血清(FBS,Adamas,中国)的RPMI-1640培养基中,置于37℃、5%二氧化碳(CO₂)的培养箱(BB150,赛默飞世尔,美国)内培养。将4T1细胞接种于培养板,培养24小时后分为4组:
1. 对照组:细胞不做任何处理;
2. BROP组:细胞与BROP共孵育,不进行其他处理;
3. 超声组:仅用1.0 MHz、1 W·cm⁻²的超声照射细胞10分钟;
4. BROP+超声组:细胞与BROP共孵育后,用1.0 MHz、1 W·cm⁻²的超声照射10分钟。
4.6 体内肿瘤抑制实验
5周龄雌性BALB/c小鼠购自上海比卡克伊生物技术有限公司,用于构建4T1肿瘤模型。将小鼠随机分为4组(n=4),在其右侧腹部皮下注射2×10⁶个4T1细胞,分组处理如下:
1. 对照组;
2. BROP组;
3. 超声组;
4. BROP+超声组。
其中,BROP组与BROP+超声组通过尾静脉注射BROP(1 mg·mL⁻¹,100 μL),超声组与BROP+超声组接受1.0 MHz、1 W·cm⁻²的超声治疗,每次10分钟。在14天的实验周期内,于第0.5天和第7.5天各处理1次,每2天测量小鼠体重与肿瘤体积,肿瘤体积计算公式如下:
公式(1)
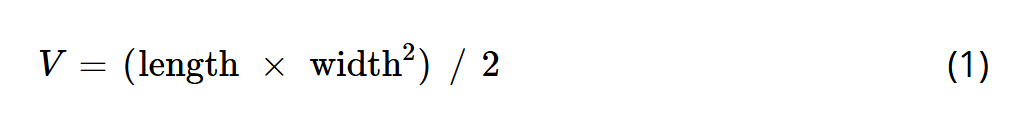
第14天时处死小鼠,剥离肿瘤组织,拍照并称重;将肿瘤组织切片后,通过苏木精-伊红(H&E)、Ki-67、末端脱氧核苷酸转移酶介导的dUTP缺口末端标记(TUNEL)、缺氧诱导因子1α(HIF-1α)及谷胱甘肽过氧化物酶4(GPX4)染色进行组织学分析。
4.7 统计分析
采用GraphPad Prism 8.0软件进行统计分析。n代表独立实验次数或每组小鼠数量,每个实验至少包含3个生物学重复,所有实验流程至少重复3次。实验数据以平均值±标准差(SD)表示:两组间比较采用双尾非配对t检验,多组间比较采用单因素方差分析(ANOVA)结合Tukey多重比较事后检验。图表中统计学显著性标注如下:ns表示无显著性差异,*表示p<0.05,**表示p<0.01,***表示p<0.001。
4.8 伦理声明
所有小鼠实验均严格遵循实验动物伦理委员会的指导原则,并经上海智北生物技术有限公司动物伦理委员会批准(批准号:IRB-AF73)。
https://wap.sciencenet.cn/blog-41174-1503109.html
上一篇:纳米气泡热酸稳定性研究
下一篇:高压氧治疗的禁忌症