ВЉЮФ
ДЯУївШЕКЫибаОПЯИНк  ОЋбЁ
ОЋбЁ
|
ЦЯЬбЬЧУєИааЭвШЕКЫидкНЕЕЭЕЭбЊЬЧЗчЯеЗНУцЕФбаОП
еЊвЊ
ЬЧФђВЁжЮСЦжаЃЌвШЕКЫиСЦЗЈв§Ц№ЕФЕЭбЊЬЧЃЈбЊЬЧЙ§ЕЭЃЉЗчЯеЙЙГЩСЫжївЊЬєеН1,2ЁЃБиаыЕїећвШЕКЫиМССПвдШЗБЃбЊЬЧжЕДІгке§ГЃЗЖЮЇФкЃЌЕЋгЩгкЖржжвђЫиЃЌШчвћЪГЕФаджЪКЭЪБМфЁЂдЫЖЏЁЂИаШОвдМАИіЬхЖдвШЕКЫиУєИаадЕФБфЛЏЃЌЪЙЕУЦЅХфвШЕКЫиМССПгыВЈЖЏЕФбЊЬЧЫЎЦНБфЕУРЇФбЁЃМДЪЙЪЧЩдЮЂИпгкЫљашМССПЕФвШЕКЫивВПЩФмЕМжТЕЭбЊЬЧЪТМўЃЌетПЩФмДгВЛЪЪЕНЩњУќЭўаВВЛЕШЁЃвђДЫЃЌГЄЦквдРДЕФФПБъвЛжБЪЧЩшМЦвЛжжЦЯЬбЬЧУєИааЭвШЕКЫиЃЌФмЙЛИљОнжмЮЇЦЯЬбЬЧЫЎЦНвдПЩФцЕФЗНЪНздЖЏЕїНкЦфЩњЮяЛюадЃЌДгЖјзюжеЪЕЯжИќКУЕФбЊЬЧПижЦЃЌЭЌЪБНЕЕЭЕЭбЊЬЧЕФЗчЯе3ЁЃдкетРяЃЌЮвУЧБЈИцСЫNNC2215ЕФЩшМЦМАЦфЬиадЃЌетЪЧвЛжжвШЕКЫиНсКЯЮяЃЌЦфЩњЮяЛюадПЩФцЕиЯьгІгкгыЬЧФђВЁЯрЙиЕФЦЯЬбЬЧЗЖЮЇЃЌетвЛЕувбдкЬхЭтКЭЬхФкЕУЕНжЄУїЁЃЭЈЙ§НЋвЛИіЦЯЬбЬЧНсКЯЕФДѓЛЗ4КЭвЛИіЦЯЬЧмеСЌНгЕНвШЕКЫиЩЯЃЌЙЄГЬЛЏСЫNNC2215ЃЌДгЖјв§ШыСЫвЛИіПЊЙиЃЌИУПЊЙиПЩвдИљОнЦЯЬбЬЧХЈЖШДђПЊКЭЙиБеЃЌДгЖјЪЙвШЕКЫидкЛюадКЭЗЧЛюадЙЙЯѓжЎМфДяЕНЦНКтЁЃЕБЦЯЬбЬЧХЈЖШДг3діМгЕН20 mMЪБЃЌNNC2215ЖдвШЕКЫиЪмЬхЕФЧзКЭСІдіМгСЫ3.2БЖЁЃдкЖЏЮябаОПжаЃЌNNC2215ЕФЦЯЬбЬЧУєИаЩњЮяЛюадБЛжЄУїПЩвдБЃЛЄУтЪмЕЭбЊЬЧЕФгАЯьЃЌЭЌЪБВПЗжИВИЧСЫЦЯЬбЬЧВЈЖЏЁЃ
жїЮФ
ЪЙгУвШЕКЫиПижЦЬЧФђВЁАщЫцзХв§ШыЕЭбЊЬЧЕФЗчЯеЃЌМДбЊЬЧжЕЕЭгк3.9 mMЃЈВЮПМЮФЯз1,2ЃЉЁЃетЪЧгЩгкаэЖрвђЫиЕМжТбЊЬЧВЈЖЏФбвддЄВтЃЌР§ШчвћЪГЕФаджЪКЭЪБМфЁЂдЫЖЏЁЂИаШОКЭИіЬхЖдвШЕКЫиУєИаадЕФБфЛЏЁЃвђДЫЃЌЬЧФђВЁЛМепБиаыЕїећЫћУЧУПЬьЕФвШЕКЫиМССПЃЈАќРЈЛљДЁКЭВЭЧАвШЕКЫиЃЉЃЌвдПМТЧетаЉвђЫиЁЃШЛЖјЃЌЮЊСЫБмУтЕЭбЊЬЧЪТМўЃЌгШЦфЪЧдквЙМфПЩФмЛсКмЮЃЯеЃЌаэЖрШЫбЁдёБЃЪиЕФвШЕКЫиМССПЁЃгЩгкКІХТЕЭбЊЬЧЖјЭзавШЕКЫиМССПЃЌЫцКѓЕМжТбЊЬЧПижЦВЛМбЃЌДгЖјдіМгСЫГЄЦкИпбЊЬЧв§ЗЂВЂЗЂжЂЕФЗчЯеЁЃЮЊСЫдкВЛдіМгЕЭбЊЬЧЗчЯеЕФЧщПіЯТДйНјИФЩЦбЊЬЧПижЦЃЌзд20ЪРМЭ70ФъДњвдРДЃЌШЫУЧвЛжБдкзЗЧѓЩшМЦвЛжжФмЙЛИљОнБфЛЏЕФбЊЬЧЫЎЦНаоИФЦфЩњЮяЛюадЕФвШЕКЫиЕФЯыЗЈ3ЁЃОЁЙмгааэЖрГіАцЮяКЭзЈРћЃЌЕЋЕНФПЧАЮЊжЙЃЌУЛгавЛжжЛњжЦБЛжЄУїФмЙЛНтОіетИіЮЪЬтЃЌвджСгкЫќПЩвдгІгУгкжЮСЦЬЧФђВЁ5,6,7,8,9,10,11ЁЃИУСьгђЕФДѓЖрЪ§ТлЮФУшЪіСЫОлКЯЮяЯЕЭГЃЌетаЉЯЕЭГПЩвдИљОнбЊЬЧВЈЖЏДгЦЄЯТЃЈs.c.ЃЉДЂПтЪЭЗХвШЕКЫиЃЌЕЋетбљЕФЯЕЭГЪмЕНбгГйЕФЦЯЬбЬЧЯђЦЄЯТРЉЩЂЕФЯожЦЃЌвдМАЪЭЗХЕФвШЕКЫиНјШыбЊвКбЛЗЕФбгГйЁЃДЫЭтЃЌетбљЕФЯЕЭГВЛПЩФцЕиЪЭЗХвШЕКЫиЃЌвтЮЖзХвЛЕЉвШЕКЫиДгДЂПтЪЭЗХЃЌЫќОЭВЛдйЪЧЦЯЬбЬЧУєИаЕФСЫЁЃвЛИіИќКУЕФЗНЗЈЫЦКѕЪЧШУвШЕКЫиБОЩэОпБИЦЯЬбЬЧЯьгІЬиадЃЌЪЙЦфФмЙЛвдПЩФцЕФЗНЪНЯьгІЦЯЬбЬЧЁЃжЕЕУзЂвтЕФЪЧЃЌЦЯЬбЬЧжЕдквЛИіЯСеЕФЗЖЮЇФкБфЛЏЃЈдкЬЧФђВЁЛМепжаДѓдМДг2ЕН20ЈC30 mMЃЉЃЌвђДЫЮЊСЫЦЯЬбЬЧУєИааЭвШЕКЫиВњЩњгАЯьЃЌБиаыдкетвЛЦЯЬбЬЧЗЖЮЇФкЛёЕУЯрЕБЖИЧЭЕФвШЕКЫиЩњЮяЛюадБфЛЏЁЃЮЊСЫДяЕНетжжЖдЦЯЬбЬЧЕФУєИаадЃЌНЋашвЊвЛИіЛЏбЇЛљЭХЃЌЫќФмЙЛдкетвЛЦЯЬбЬЧЗЖЮЇФквдзюДѓСщУєЖШгыЦЯЬбЬЧНсКЯЁЃвЛИіЯЕЭГЛљгкЙбОлбвдхЬЧ/ИЪТЖЬЧвШЕКЫиНсКЯЮяЃЌЫќУЧПЩвддкЦЯЬбЬЧУєИаЕигыИЪТЖЬЧЪмЬхНсКЯгывШЕКЫигывШЕКЫиЪмЬхНсКЯжЎМфЕФЦНКтжаДгбЛЗжаЧхГ§12ЃЌЕЋетВЂУЛгаГЌЙ§IЦкСйДВЪдбщОЭжЕЕУНјвЛВНзЗЧѓ13ЁЃЗЂЯжЦЯЬбЬЧЗДгІЪЧЧГЕФЃЌИЪТЖЬЧЪмЬхЕФИпЧхГ§ТЪЕМжТЗЧГЃЕЭЕФЬхФкаЇФмЃЌетвтЮЖзХзюжеашвЊНћжЙИпЕФвШЕКЫиМССПЁЃ
ЖрФъРДЃЌШЫУЧвЛжБдкЬНЫїНЋЦЯЬбЬЧУєИаПЊЙив§ШывШЕКЫиЗжзгжаЕФИХФю1,14,15,16ЁЃвЛИіПЊЙиЩцМАдквШЕКЫиЩЯЫЋжиНсКЯвЛИіЦЯЬбЬЧНсКЯЛљађКЭвЛИіНсКЯАщТТЃЌетбљдкЕЭЦЯЬбЬЧХЈЖШЯТЃЌПЊЙиЛсгеЕМвЛИіБеКЯЕФЕЭЛюадзДЬЌЃЌЫцзХЦЯЬбЬЧХЈЖШЕФдіМгЃЌЦНКтЛсЯђвЛИіПЊЗХЕФИпЛюадзДЬЌЧуаБЁЃвђДЫЃЌЦЯЬбЬЧНсКЯЛљађБиаыдкЬЧФђВЁЛМепЬхФкГіЯжЕФЯСеЦЯЬбЬЧЗЖЮЇЃЈдМ2ЕН20ЈC30 mMЃЉФкЭЌЪБЖдЦЯЬбЬЧКЭНсКЯАщТТОпгаЧзКЭСІЁЃДЫЭтЃЌПЊЙиЕФСНИізщГЩВПЗжБиаывдФГжжЗНЪНСЌНгЕНвШЕКЫиЩЯЃЌвдШЗБЃдкБеКЯзДЬЌЯТЃЌЭЈЙ§ИФБфвШЕКЫиЙЙЯѓКЭ/ЛђзшЖЯвШЕКЫиЕФЪмЬхНсКЯБэУцЃЌНЕЕЭвШЕКЫиЕФЩњЮяЛюадЁЃетИіЯыЗЈвбОЭЈЙ§ЪЙгУХ№ЫсѕЅзїЮЊЦЯЬбЬЧНсКЯМСРДзЗЧѓЃЌЕЋЦљНёЮЊжЙЃЌДЫРрЩшМЦЕФЦЯЬбЬЧУєИаадЖдгквЉРэгУЭОРДЫЕЛЙЬЋгаЯоЁЃжЎЧАЙигкгывШЕКЫивЛЦ№ЙЄзїЕФЬМЫЎЛЏКЯЮяУєИаПЊЙиЕФзюМбЪОР§ЯдЪОСЫЖдИпХЈЖШЙћЬЧЃЈ50 mMЃЉЕФУєИаадЃЌЕЋИУЛЏКЯЮяЖдЦЯЬбЬЧВЛУєИа15ЁЃзюНќЗЂЯжЕФвЛжжДѓЛЗЬсЙЉСЫСэвЛжжЦЯЬбЬЧНсКЯдЊЫиЕФбЁдё4ЁЃЩшМЦетжжДѓЛЗЪЧЮЊСЫЬсЙЉвЛИіЦЯЬбЬЧНсКЯЧЛЪвЃЌИУЧЛЪвШЗБЃЖдЦЯЬбЬЧЕФЯрЙиЧзКЭСІвдМАЯрЖдгкЦфЫћЬМЫЎЛЏКЯЮяКЭПЩФмИЩШХЕФаЁЗжзгЕФбЁдёадЁЃдкетРяЃЌЮвУЧУшЪіСЫNNC2215ЕФЗжзгЩшМЦЃЌетЪЧвЛжжЭЈЙ§дкB29LysДІв§ШыДѓЛЗВЂдкB1PheДІЭЈЙ§ЖЬСЌНгзгв§ШыO1-ЦЯЬЧмеЖјОпгаЦЯЬбЬЧПЊЙиЕФвШЕКЫиЃЈЭМ1aЃЉЁЃетжжЦЯЬбЬЧНсКЯМСЁЂЦЯЬЧмеЁЂСЌНгзгКЭХМСЊЮЛЕуЕФзщКЯБЛжЄУїИГгшСЫNNC2215ЦЯЬбЬЧУєИаЕФЩњЮяЛюадЃЌЕБЦЯЬбЬЧДг0діМгЕН20 mMЪБЃЌвШЕКЫиЪмЬхНсКЯЧзКЭСІдіМгСЫ12.5БЖЃЌДг3діМгЕН20 mMЪБдіМгСЫ3.2БЖЁЃДЫЭтЃЌNNC2215БЛжЄУїдкЬхФкЖдЦЯЬбЬЧУєИаЃЌФмЙЛМѕЧсжэЕФЕЭбЊЬЧЃЌВЂМѕЩйЬЧФђВЁДѓЪѓПкЗўЦЯЬбЬЧФЭСПЪдбщЃЈGTTsЃЉЦкМфЕФЦЯЬбЬЧВЈЖЏЁЃ
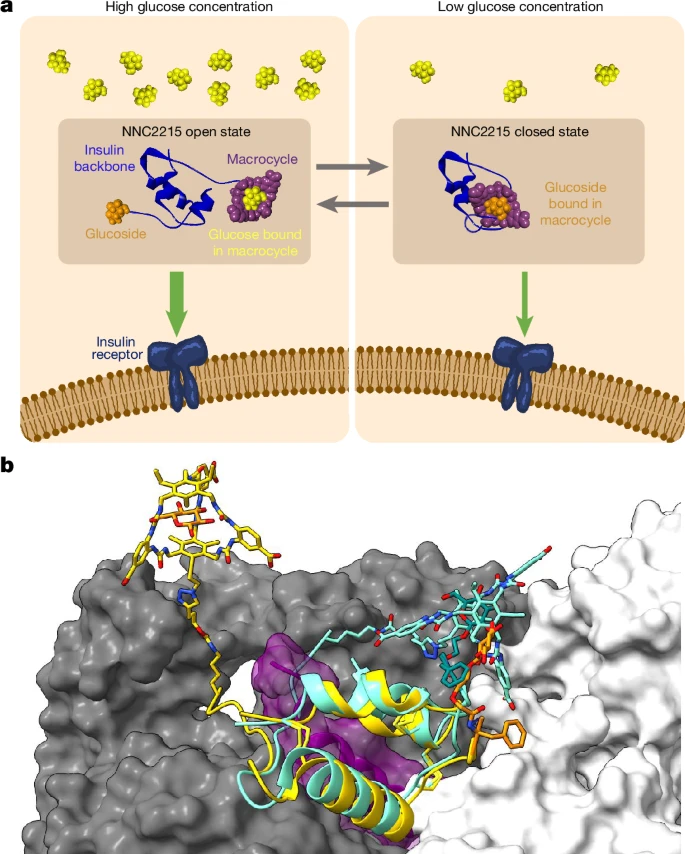
ЭМ1ЃКNNC2215ЕФЙІФмдРэКЭ3DФЃаЭЁЃ
aЃЌNNC2215ЪЧвЛИіОпгаЦЯЬбЬЧУєИаПЊЙиЕФвШЕКЫиЗжзгЁЃЫцзХЦЯЬбЬЧХЈЖШЕФдіМгЃЌПЊЙиЦНКтГЏЯђПЊЗХзДЬЌЃЌNNC2215ЕФвШЕКЫиЪмЬхЧзКЭСІИпЃЌДгЖјгажњгкЗРжЙИпбЊЬЧЁЃЕБЦЯЬбЬЧЫЎЦНЯТНЕЪБЃЌПЊЙиЦНКтГЏЯђБеКЯзДЬЌЃЌИЩШХNNC2215гывШЕКЫиЪмЬхЕФНсКЯФмСІЃЌДгЖјгажњгкЗРжЙЕЭбЊЬЧЁЃвШЕКЫижїСДЁЂДѓЛЗЁЂЦЯЬЧмеКЭЦЯЬбЬЧФЃаЭЪЧЪЙгУBIOVIA Discovery StudioЃЈДяЫїЯЕЭГЙЋЫОЃЉжЦБИЕФЁЃbЃЌNNC2215дкПЊЗХКЭБеКЯаЮЪНЕФ3DФЃаЭЁЃвШЕКЫижїСДЯдЪОЮЊДјзДЃЌПЊЙидЊМўЃЈЦЯЬЧмеКЭДѓЛЗЃЉЯдЪОЮЊЙїзДБэЪОЁЃРДздPDB 6PXVЕФвШЕКЫиЪмЬхСДAКЭCЯдЪОЮЊАзЩЋКЭЛвЩЋЕФБэУцБэЪОЁЃNNC2215ЕФПЊЗХаЮЪНЃЈЛЦЩЋЃЉгаздгЩЕФЦЯЬбЬЧЃЈГШЩЋЃЛзѓЩЯНЧЃЉАѓЖЈдкДѓЛЗЩЯB29ДІЁЃNNC2215ПЊЗХаЮЪНЕФB1ЈCЦЯЬЧмеЯдЪОЮЊГШЩЋЃЈгвВрЃЉЁЃNNC2215ЕФБеКЯаЮЪНЃЈЧрЩЋЃЉгаЦЯЬЧмеАѓЖЈдкДѓЛЗжаЃЌВЂЯдЪОвШЕКЫиBСДЕФCЖЫВПЗжЃЈАќРЈПЊЙиЃЉгывШЕКЫиЪмЬхЕФCЖЫгђЃЈІС-CTЃЌзЯЩЋЃЉжЎМфЕФГхЭЛЁЃ
ЛЏбЇ
ЭЈЙ§дкШ§пђаЮГЩЙ§ГЬжаНЋДјгаЕўЕЊБћЛљСЌНгзгЕФДѓЛЗбмЩњЮягыB29Lys NІХАБЛљзщСЌНгЃЌНЋДѓЛЗ4ЃЈЭМ2aЃЉХМСЊЕНdesB30ШЫвШЕКЫиЩЯ17ЁЃЭЈЙ§етжжЗНЪНЃЌБмУтСЫашвЊе§НЛЕиДІРэШ§ИієШЫсжЎвЛЕФашЧѓЃЌетЖдгкСЌНгЕНДѓЛЗжЇжљЩЯЕФШЮКЮвЛИіCOOHЛљЭХРДЫЕЪЧБиашЕФЁЃзюГѕБЈЕРЕФДѓЛЗЪЙгУЕФЪїжІзДОлКЯЮядкгывШЕКЫи-ДѓЛЗХМСЊЮявЛЦ№ЙЄзїЪББЛЗЂЯжЪЧВЛБивЊЕФЁЃГ§СЫB29ДІЕФДѓЛЗЭтЃЌЭЈЙ§дкpH 7.5ЯТЪЙгУЯргІЕФфхДњШ§ЗњМзЛљЛЧЫсБНѕЅЃЌНЋДјгаЖЬСЌНгзгЕФO1-ЦЯЬЧмеИНзХЕНBСДNЖЫАБЛљзщЃЈPheB1ЃЉЩЯЃЈВЮПМЮФЯз18ЃЉЁЃЦЯЬЧмевдЦфO-ШЋввѕЃБЃЛЄЕФНЈжўПщаЮЪНЪЙгУЃЌВЂЭЈЙ§ЮТКЭдэЛЏДгвШЕКЫиХМСЊЮяжаШЅГ§СЫввѕЃЛљЁЃЭЈЙ§ЪЁТдЦЯЬЧмеВНжшЃЌвдРрЫЦNNC2215ЕФЗНЪНжЦБИСЫНідкB29ДІгаДѓЛЗЕФЖдееЛЏКЯЮяNNC2215aЁЃЭЈЙ§вКЯрЩЋЦз-жЪЦзСЊгУЃЈLCЈCMSЃЉЗжЮіОЙ§вШЕААзУИДІРэШЛКѓНјааШ§(2-єШввЛљ)ьЂЃЈTCEPЃЉДІРэЕФбљЦЗЃЌМЧТМСЫвШЕКЫиЩЯЕФХМСЊЮЛЕуЁЃвШЕААзУИДІРэЧаИюСЫNNC2215ЕФB22ArgжЎКѓЃЌЪЭЗХСЫB23ЈCB29ЦЌЖЮЃЌTCEPЧаЖЯСЫNNC2215ЕФЖўСђМќЃЌЗжРыГіAКЭBСДЁЃLCЈCMSЗжЮіЫљЕУЕФЗжЮіЛьКЯЮяЯдЪОДѓЛЗИНзХдкB23ЈCB29ЦЌЖЮЩЯЃЌЦЯЬЧмеИНзХдкB1ЈCB22ЦЌЖЮЩЯЃЌвдМАгЮРыЕФAСДЃЈЛЏбЇЯИНкКЭLCЈCMSЙтЦзЬсЙЉдкВЙГфаХЯЂКЭВЙГфЪ§ОнжаЃЉЁЃ
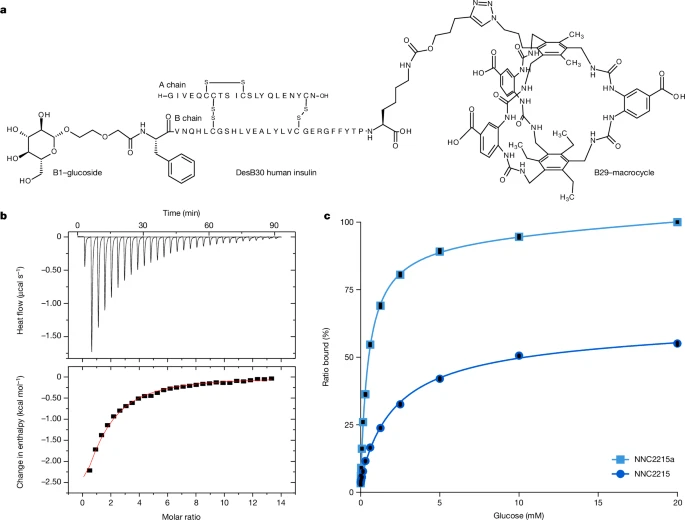
ЭМ2ЃКNNC2215ЕФЛЏбЇНсЙЙКЭЦЯЬбЬЧНсКЯЬиад
aЃЌNNC2215дкB29КЭB1ЗжБ№СЌНгДѓЛЗКЭЦЯЬЧмеЁЃЖдееЛЏКЯЮяNNC2215aжЛдкB29СЌНгСЫДѓЛЗЁЃЪЙгУBIOVIA DrawЃЈDassault SystЈЈmesЃЉзМБИЁЃbЃЌЭЈЙ§ЕШЮТЕЮЖЈСПШШЗЈЃЈITCЃЉВтСПДѓЛЗгыЦЯЬбЬЧЕФНсКЯЧзКЭСІЁЃЭЈЙ§НЋЪ§ОнФтКЯЕН1:1НсКЯФЃаЭЃЌЛёЕУСЫ98 µMЕФKdжЕЁЃcЃЌРћгУжЪЦзЃЈMSЃЉЗжЮібаОПNNC2215ЖдЦЯЬбЬЧЕФНсКЯЁЃШчдЄЦкЕФФЧбљЃЌЫЋСЌНгЮяжаЦЯЬЧмеЕФДцдкИФБфСЫNNC2215ЯрЖдгкздгЩДѓЛЗЕФЦЯЬбЬЧЧзКЭСІЃЌЪЙЕУNNC2215гыЦЯЬбЬЧНсКЯЕФKdЮЊ2.1 mMЃЌетдкЬЧФђВЁЕФБГОАЯТБЛШЯЮЊЪЧбЯжиЕФЕЭбЊЬЧЁЃЖдееЛЏКЯЮяNNC2215aжЛАќКЌДѓЛЗЃЌЭЈЙ§жЪЦзЗЂЯжЦфгыЦЯЬбЬЧНсКЯЕФKdЮЊ0.5 mMЁЃПЩвддЄМћЃЌНЋДѓЛЗСЌНгЕНвШЕКЫиЩЯЛсЪЪЖШИФБфЦфЯрЖдгкздгЩДѓЛЗЕФЦЯЬбЬЧЧзКЭСІЁЃЭЌбљЕиЃЌНЋАзЕААзНсКЯВПЗжСЌНгЕНвШЕКЫиЛђЦфЫћЕААзжЪЩЯвВЭЈГЃЛсИФБфЫќУЧЯрЖдгкздгЩХфЬхЕФЧзКЭСІЃЌетПЩФмЪЧгЩгкСЌНгВПЗжгыСкНќЕААзжЪМАЦфВрСДЕФШѕЯрЛЅзїгУЫљжТЁЃзмЬхЖјбдЃЌЭЈЙ§жЪЦзВтЕУЕФNNC2215КЭЖдееЛЏКЯЮяNNC2215aЕФЦЯЬбЬЧЧзКЭСІБШздгЩДѓЛЗЗжБ№МѕШѕСЫдМ20БЖКЭ5БЖЃЌДгЖјжЄУїСЫКЯЪЪЕФПЊЙиЖЏЬЌвбОЪЕЯжЁЃNNC2215КЭNNC2215aЯьгІ0жС20 mMЦЯЬбЬЧЕФЦЯЬбЬЧНсКЯКЭЭЌЪБЗЂЩњЕФПЊЙиДђПЊПЩвдЭЈЙ§ЭМ2cжаЕФжЪЦзНсКЯЭМНјааИњзйЁЃНсКЯЧњЯпЕФзюЖИВПЗжгыБЛШЯЮЊЪЧЕЭбЊЬЧЕФЧщПівЛжТЃЌМДЦЯЬбЬЧХЈЖШЕЭгк4 mMЁЃОЁЙмжЪЦзЪ§ОнЪЧдкЦјЯрЃЈжЪЦзецПеЃЉжаЛёЕУЕФЃЌЕЋетаЉЪ§ОнЭЈГЃЗДгГСЫДгжаШЁбљЕФШмвКжаЗжзгЕФЯрЛЅзїгУ19ЁЃ
NNC2215ЈCвШЕКЫиЪмЬхЕФ3DФЃаЭ
ЭЈЙ§НЋNNC2215ЕФФЃаЭЕўМгЕНвШЕКЫи-вШЕКЫиЪмЬхИДКЯЮяЕФЕААзжЪЪ§ОнПтЃЈPDBЃЉ6PXVЃЈВЮПМЮФЯз20,21,22,23,24ЃЉЩЯРДЙЙНЈНсЙЙФЃаЭЃЌвдЪОЗЖЦфЙІФмЁЃШчЭМ1bЫљЪОЃЌЕБПЊЙиБеКЯЪБЃЌвШЕКЫиBСДЕФCЖЫВПЗжгывШЕКЫиЪмЬхЕФCЖЫНсЙЙгђЃЈГЦЮЊІС-CTЃЉжЎМфЗЂЩњГхЭЛЁЃІС-CTНсЙЙгђЪЧвШЕКЫиНсКЯЕФЙиМќВПЗж25ЁЃЮвУЧШЯЮЊЃЌетжжПеМфЮЛзшЪЧЙлВьЕНЕФNNC2215дкЕЭЦЯЬбЬЧХЈЖШЯТЃЈМДПЊЙижївЊДІгкБеКЯЙЙЯѓЪБЃЉЪмЬхЧзКЭСІНЕЕЭЕФЧ§ЖЏСІЁЃЯрБШжЎЯТЃЌПЊЗХзДЬЌЕФФЃаЭгыФмЙЛгыЪмЬхНсКЯЕФЭъШЋЛюадвШЕКЫиЙЙЯѓМцШнЁЃ
ЬхЭтбаОП
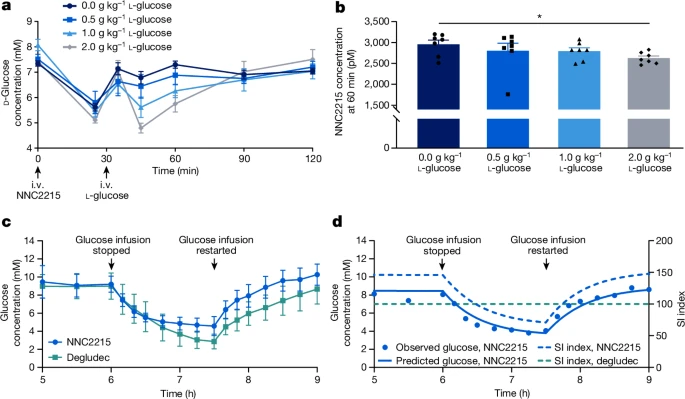
ЮЊСЫбаОПNNC2215гывШЕКЫиЪмЬхЕФЦЯЬбЬЧУєИаЯрЛЅзїгУЃЌНјааСЫгаЮоВЛЭЌЦЯЬбЬЧХЈЖШЯТЕФвШЕКЫиЪмЬхНсКЯбаОПЃЌЦфжаБШНЯСЫШЫвШЕКЫиЪмЬхAЃЈhIR-AЃЉгыNNC2215ЁЂШЫвШЕКЫиКЭвШЕКЫиЕТЙШСаЭЁЃЈвЛжжѕЃЛЏЕФГЄаЇЛљДЁвШЕКЫиЃЉЕФНсКЯЃЈЭМ3aЈCcЃЉЁЃЖРЬижЎДІдкгкЃЌПЩвдПДЕНЫцзХЦЯЬбЬЧХЈЖШЕФдіМгЃЌNNC2215ЕФНсКЯЧњЯпЪмЕНгАЯьЃЈЭМ3aЃЉЁЃЯрЖдгкhIR-AЖдШЫвШЕКЫиЕФЧзКЭСІЃЌNNC2215ЕФЧзКЭСІДг0діМгЕН20 mMЦЯЬбЬЧЪБЕФ0.75%діМгЕН2.9%, 4.3%, 6.5%КЭ9.2%ЃЌЖјвШЕКЫиЕТЙШСаЭЁЯрЖдгкШЫвШЕКЫиОпгаКуЖЈЕФЧзКЭСІЃЈБэ1КЭЭМ3dЃЉЁЃДг0ЕН20 mMЦЯЬбЬЧЃЌhIR-AЖдNNC2215ЕФЧзКЭСІдіМгСЫ12.5БЖЃЌЖјДг3ЕН20 mMЦЯЬбЬЧдђдіМгСЫ3.2БЖЃЌетЪЧЬЧФђВЁЛМепжаПЩЙлВьЕНЕФХЈЖШЗЖЮЇЃЈБэ1ЃЉЁЃжЕЕУзЂвтЕФЪЧЃЌетИідіМгЕФhIR-AЧзКЭСІЪЧдкДцдк1.5%АзЕААзЕФЧщПіЯТШЗЖЈЕФЃЌетЪЧИУВтЖЈжаФмШнШЬЕФзюДѓХЈЖШЃЈгыШЫЬхФкДѓдМ4%ЕФАзЕААзЯрБШЃЉЁЃдкУЛгаАзЕААзЕФЧщПіЯТЃЌДг0ЕН20 mMЦЯЬбЬЧЃЌhIR-AЖдNNC2215ЕФЧзКЭСІдіМгМѕЩйЕН6.8БЖЃЈЭМРЉеЙЪ§ОнЭМ1aЃЉЁЃДЫЭтЃЌЙлВьЕНдкДцдк1.5%АзЕААзЕФЧщПіЯТЃЌЖдгкНіКЌДѓЛЗЕФЖдееЛЏКЯЮяNNC2215aЃЌhIR-AЕФЧзКЭСІвВгааЁЗљЖШЕФдіМгЃЈДг0ЕН20 mMЦЯЬбЬЧдіМгСЫ2.4БЖЃЉЃЌетвВБэУїСЫШѕЕФАзЕААзНсКЯаЇЙћЃЈЭМРЉеЙЪ§ОнЭМ1bЃЉЁЃ
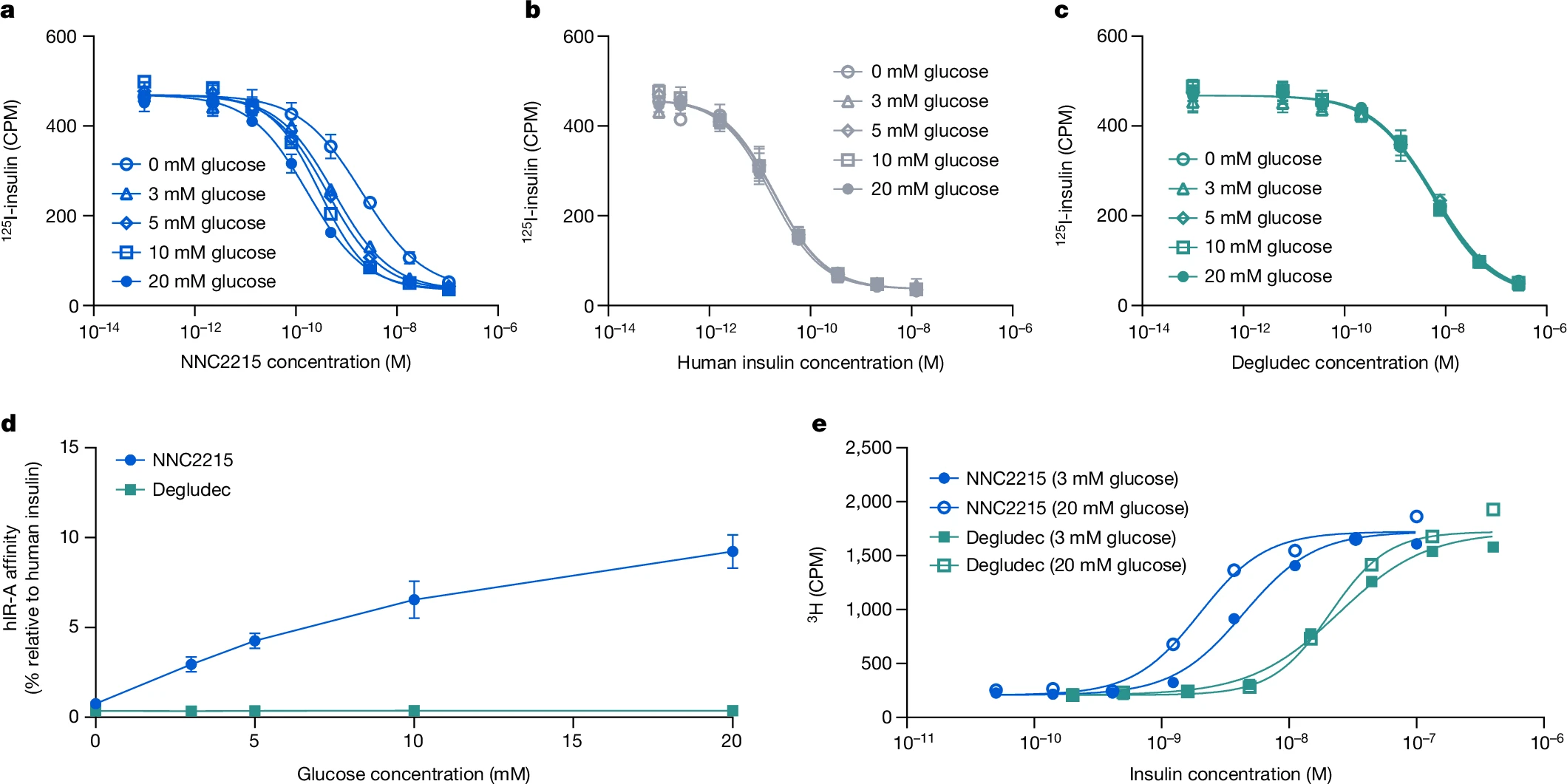
ЭМ3ЃКNNC2215ЕФhIR-AЧзКЭСІгыЦЯЬбЬЧХЈЖШЕФЙиЯЕ
aЈCcЃЌдк0жС20 mM d-ЦЯЬбЬЧДцдкЯТЃЌNNC2215ЃЈaЃЉЁЂШЫРрвШЕКЫиЃЈbЃЉКЭвШЕКЫиЕТЙШЃЈcЃЉДгhIR-AжУЛЛ125I-вШЕКЫиЕФДњБэадЧњЯпЁЃЪ§ОнЮЊОљжЕЁРБъзМВюЁЃn=3ДЮММЪѕжиИДЁЃЖдгкФГаЉЪ§ОнЕуЃЌБъзМВюЮѓВюЬѕаЁгкЗћКХДѓаЁЁЃdЃЌдкВЛЭЌЦЯЬбЬЧХЈЖШЯТЃЌNNC2215КЭвШЕКЫиЕТЙШЯрЖдгкШЫРрвШЕКЫиЕФhIR-AЧзКЭСІЁЃЪ§ОнЮЊОљжЕЁРБъзМВюЁЃn=3ДЮЖРСЂжиИДЁЃЖдгкФГаЉЪ§ОнЕуЃЌБъзМВюЮѓВюЬѕаЁгкЗћКХДѓаЁЁЃeЃЌдкЕЭЃЈ3 mMЃЉКЭИпЃЈ20 mMЃЉl-ЦЯЬбЬЧХЈЖШЯТЃЌNNC2215КЭвШЕКЫиЕТЙШМССПвРРЕадЕиНЋ3H-d-ЦЯЬбЬЧзЊЛЏЮЊжЌжЪЕФДњБэадЧњЯпЁЃЪ§ОнЮЊОљжЕЁЃn=2ДЮММЪѕжиИДЁЃCPMЃЌУПЗжжгМЦЪ§ЁЃ
Бэ1 NNC2215ЛюадЖдhIR-AНсКЯКЭЯТгЮДњаЛЗДгІЕФЦЯЬбЬЧУєИаад
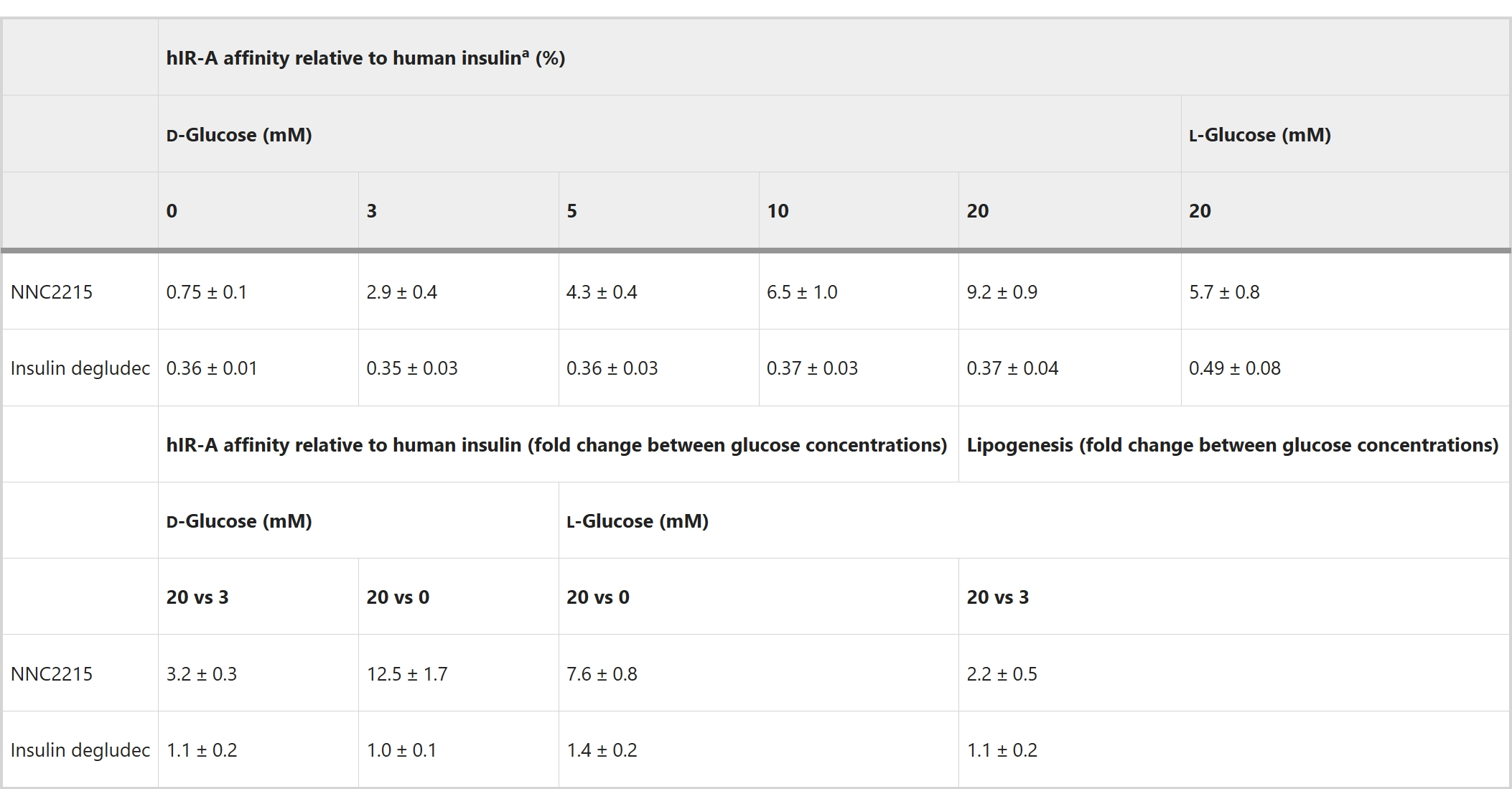
гывШЕКЫибљЩњГЄвђзг1ЪмЬхЃЈIGF-1RЃЉЯрБШЃЌЖдвШЕКЫиЪмЬхЕФЬивьадЗЧГЃживЊЃЌвдБмУтШЮКЮдіМгЕФДйгаЫПЗжСбЛюадЁЃдкгаЮо20 mM d-ЦЯЬбЬЧЕФЧщПіЯТВтСПСЫIGF-1RЕФНсКЯЃЈРЉеЙЪ§ОнЭМ1cЃЉЁЃЯрЖдгкШЫРрвШЕКЫиЃЌдк0КЭ20 mMЦЯЬбЬЧЪБЃЌNNC2215ЕФIGF-1RЧзКЭСІдМЮЊЦфвШЕКЫиЪмЬхЧзКЭСІЕФ10%ЃЈРЉеЙЪ§ОнЭМ1aЁЂdЃЉЁЃвђДЫЃЌгыШЫРрвШЕКЫиЯрБШЃЌNNC2215ЖдвШЕКЫиЪмЬхЕФЬивьадИпгкЖдIGF-1RЕФЬивьадЁЃ
дкБэДяПЫТЁШЫРрвШЕКЫиЪмЬхЕФCHO-hIRЯИАћжаМьВщСЫNNC2215МЄЛювШЕКЫиаХКХЭЈТЗЕФФмСІЁЃЛёЕУСЫNNC2215ДЬМЄШЫРрвШЕКЫиЪмЬхРвАБЫсСзЫсЛЏЕФЭъећМССП-ЗДгІЧњЯпЃЌЦфаЇФмЮЊ57.1%ЃЈ95%жУаХЧјМфЃЈCIЃЉ=41.7ЈC78.1ЃЉЁЃЭЈЙ§AKTКЭERKМЄЛюЕФЯТгЮаХКХДЋЕМЯрЖдгкШЫРрвШЕКЫиОпгаЯрЭЌЕФЦНКтЃЌаЇФмЗжБ№ЮЊ68.4%ЃЈ95% CI=55.7ЈC84.0ЃЉКЭ48.0%ЃЈ95% CI=34.4ЈC66.9ЃЉЃЈРЉеЙЪ§ОнЭМ1eЈCgЃЉЁЃ
NNC2215ЕФЦЯЬбЬЧУєИаадЗДгГдкДѓЪѓжЌЗОЯИАћжаВтСПЕФЬхЭтДњаЛжеЕуЩЯЁЃдкРћгУЩњРэЛюадd-ЦЯЬбЬЧЕФЖдгГЬхЃЌМДl-ЦЯЬбЬЧЕФгХЪЦЯТЃЌЮвУЧВтСПСЫДѓЪѓжЌЗОЯИАћжаЕФЦЯЬбЬЧЩуШЁКЭВЂШыжЌжЪЕФЙ§ГЬЃЌМДЩњжЌзїгУЁЃЪЙгУl-ЦЯЬбЬЧЕФРэгЩЪЧЃЌжСЩйдкФГаЉЬѕМўЯТЃЌЯИАћВЛЛсвдПЩБЅКЭЕФЗНЪНДпЛЏЛђЩуШЁl-ЦЯЬбЬЧЃЌГ§СЫвЛаЉИяРМЪЯвѕадОњКЭжВЮяЭтЁЃдкЖдееЪЕбщжаЃЌl-ЦЯЬбЬЧВЛгыd-ЦЯЬбЬЧОКељНјШыжЌЗОЯИАћЕФЩуШЁЃЌЕЋl-ЦЯЬбЬЧгыNNC2215ЕФЗЧЪжадДѓЛЗНсКЯЕФЧзКЭСІгыd-ЦЯЬбЬЧЯрЭЌЁЃдкhIR-AНсКЯВтЖЈжаЃЌl-ЦЯЬбЬЧдкМЄЛюNNC2215ЗНУцЕФаЇСІВЛШчd-ЦЯЬбЬЧЁЃдк0жС20 mMжЎМфЙлВьЕНЕФNNC2215НсКЯЧзКЭСІЕФдіМгЖдгкl-ЦЯЬбЬЧЪЧ7.6БЖЃЌЖјЖдгкd-ЦЯЬбЬЧЪЧ12.5БЖЁЃвђДЫЃЌгУl-ЦЯЬбЬЧВтЕУЕФДњаЛЗДгІНЋЕЭгкгУd-ЦЯЬбЬЧЕФаЇЙћЁЃЩњжЌзїгУВтСПЯдЪОЃЌдкЕЭЃЈ3 mMЃЉгыИпЃЈ20 mMЃЉl-ЦЯЬбЬЧЯТЃЌгЩNNC2215геЕМЕФ3H-d-ЦЯЬбЬЧзЊЛЏЮЊжЌжЪЕФАызюДѓаЇгІМССПВювьЮЊ2.2БЖЃЈЭМ3eЃЉЁЃетжЄУїСЫNNC2215дкЬхЭтЙигквШЕКЫигеЕМЕФДњаЛЗДгІЕФЦЯЬбЬЧУєИаадЁЃЪЙгУвШЕКЫиЕТЙШЪБЃЌЩњжЌзїгУЖдl-ЦЯЬбЬЧЮоЯьгІЁЃ
ЬхФквЉРэбЇ
ЮЊСЫбаОПNNC2215дкЬхФкЕФЦЯЬбЬЧХЈЖШУєИаЕФМЄЛюКЭЪЇЛюЃЌЮвУЧПЊЗЂСЫШ§жжВЛЭЌЕФЗНАИЁЃзюМђЕЅЕФЗНАИЪЧЃЌДѓЪѓОВТізЂЩфNNC2215КѓИјгшl-ЦЯЬбЬЧЃЌДгЖјДЅЗЂNNC2215ЃЌЕМжТd-ЦЯЬбЬЧНЕЕЭЃЌЧветжжНЕЕЭвРРЕгкl-ЦЯЬбЬЧЕФМССПЁЃДЫЭтЃЌдкжэжаЭЈЙ§МБадНЕЕЭбЊНЌЦЯЬбЬЧРДбаОПNNC2215дкЕЭЦЯЬбЬЧЯТЕФЪЇЛюЃЌгыЗЧЦЯЬбЬЧУєИавШЕКЫиЃЈвШЕКЫиЕТЙШЃЉв§Ц№ЕФбЊЬЧЯТНЕНјааБШНЯЁЃзюКѓЃЌдкЬЧФђВЁДѓЪѓжаНјааЦЯЬбЬЧЬєеНЦкМфЃЌбаОПСЫNNC2215дкРрЫЦВЭКѓЦЯЬбЬЧВЈЖЏЦкМфЕФЛюЛЏЁЃгЩгкРрЫЦЕФвЉДњЖЏСІбЇЬиадЃЌвШЕКЫиЕТЙШБЛгУзїОВТізЂЩфДѓЪѓбаОПЕФЖдеезщКЭжэбаОПжаЕФБШНЯзщЃЈРЉеЙЪ§ОнБэ1aЃЉЃЌЖјдкДѓЪѓЦЯЬбЬЧЬєеНбаОПжаЃЌШЫРрвШЕКЫиБЛгУзїЖдеезщЁЃЭЈЙ§ИјДѓЪѓОВТізЂЩфШЗЖЈNNC2215ЕФЬхФкАыЫЅЦкЮЊ1.2аЁЪБЃЌИјжэОВТізЂЩфШЗЖЈЮЊ1.3аЁЪБЃЈРЉеЙЪ§ОнБэ1aЃЉЁЃ
ЭМ4: NNC2215дкЬхФкЕФЦЯЬбЬЧХЈЖШУєИаадМЄЛюКЭЪЇЛю
ЭМ4aЭЈЙ§ОВТізЂЩфВЛЭЌМССПЕФNNC2215ВЂЫцКѓИјгшl-ЦЯЬбЬЧЃЈАќРЈдиЬхЃЉРДДЅЗЂNNC2215ЕФвШЕКЫизїгУЃЌЕМжТd-ЦЯЬбЬЧНЕЕЭЁЃЪ§ОнЮЊУПзщ7жЛЖЏЮяЕФЦНОљжЕЁРБъзМЮѓЁЃ
ЭМ4bЭЈЙ§ВтСП60ЗжжгКѓЕФNNC2215ХЈЖШЦРЙРNNC2215ЕФЦЯЬбЬЧМССПвРРЕадЧхГ§ЁЃЪ§ОнЮЊУПзщ7жЛЖЏЮяЕФЦНОљжЕЁРБъзМЮѓЁЃЭГМЦЗжЮіВЩгУЫЋВрбЇЩњtМьбщЃЛ*P=0.013ЁЃ
ЭМ4cдкЭЃжЙВЂжиаТЦєЖЏКуЖЈОВТіЪфзЂЦЯЬбЬЧЃЈ6КСПЫ/ЧЇПЫ/ЗжжгЃЉКѓЃЌГжајОВТіЪфзЂNNC2215ЃЈ1.86ЦЄФІЖћ/ЧЇПЫ/ЗжжгЃЉЛђвШЕКЫиЕТЙШСаЃЈ0.9ЦЄФІЖћ/ЧЇПЫ/ЗжжгЃЉЦкМфЃЌLYDжэЕФДњБэадбЊЬЧТжРЊЁЃЪ§ОнЮЊУПзщ7жЛЃЈNNC2215ЃЉКЭ8жЛЃЈвШЕКЫиЕТЙШСаЃЉжэЕФЦНОљжЕЁРБъзМВюЁЃ
ЭМ4dЖдвЛжЛНгЪм1.72ЦЄФІЖћ/ЧЇПЫ/ЗжжгNNC2215МССПЕФLYDжэНјаавЉДњЖЏСІбЇ/вЉаЇбЇНЈФЃЕФНсЙћЁЃSIЃЌвШЕКЫиУєИааджИЪ§ЁЃ
ЕЭбЊЬЧбаОПдкLYDжэжа
ЮЊСЫЦРЙРНЁПЕLandraceЈCYorkshireЈCDurocЃЈLYDЃЉжэжаNNC2215ЕФЬхФкЛюадЃЌВЂЛљБОЩЯЯћГ§d-ЦЯЬбЬЧВЈЖЏЖдФкдДадМЄЫиЪЭЗХЕФгАЯьЃЌЪЙгУСЫЩњГЄвжЫиЪфзЂРДвжжЦвШИпбЊЬЧЫиКЭвШЕКЫиЕФЗжУкЁЃДЫЭтЃЌИУбаОПдкВЛЭЌЕФЪфзЂЫйТЪЯТЃЌАщЫцвШИпбЊЬЧЫиЬцДњКЭКуЖЈЦЯЬбЬЧЪфзЂЃЌНјааСЫNNC2215гывШЕКЫиЕТЙШСаЕФБШНЯЁЃ5аЁЪБЪфзЂКѓЃЌЕБДяЕНвШЕКЫиЃЈЕТЙШСаЛђNNC2215ЃЉКЭЦЯЬбЬЧЕФНќЫЦЮШЬЌбЊНЌХЈЖШЪБЃЌЭЃжЙd-ЦЯЬбЬЧЪфзЂЃЈМћЭМ4cЃЉЁЃДЫГЬађЕМжТбЊНЌd-ЦЯЬбЬЧЯТНЕЃЌгЩГжајЕФвШЕКЫиЪфзЂДйНјЃЌЕЋгывШЕКЫиЕТЙШСаЯрБШЃЌNNC2215ЕФЯТНЕНЯЩйЃЌДгЖјЯдЪОГівШЕКЫиЩњЮяЛюадЕФЦЯЬбЬЧУєИаЧаЛЛЁЃЪЙгУВЛЭЌЕФвШЕКЫиЪфзЂЫйТЪЦРЙРЭъећЪ§ОнМЏБэУїЃЌдкЭЃжЙЦЯЬбЬЧЪфзЂКѓЃЌЖдгкШЮКЮИјЖЈЕФбЊНЌЦЯЬбЬЧХЈЖШЃЌNNC2215ЕФбЊНЌЦЯЬбЬЧЯТНЕЗљЖШаЁгквШЕКЫиЕТЙШСаЃЈРЉеЙЪ§ОнЭМ3ЃЉЁЃдкCыФЗДгІжаЕФДѓБъзМВюЪЧгЩгквЛИідкЫљгаСљЬьЪЕбщЦкМфCыФЫЎЦННЯИпЕФЖЏЮяв§Ц№ЕФЃЈРЉеЙЪ§ОнЭМ3dЃЉЁЃЪЁТдетИіЖЏЮяЕФЪ§ОнВЂВЛИФБфЙигкЦЯЬбЬЧУєИаадЕФНсТлЃЈРЉеЙЪ§ОнЭМ3cЃЉЁЃЭМ4cжаЯдЪОСЫСНИіДњБэадзщЕФЪ§ОнЃЌЦфжаЦЯЬбЬЧдкЪЙгУNNC2215ЪБЯТНЕЕНдМ4.5КСФІЖћ/Щ§ЃЌЖјдкЪЙгУвШЕКЫиЕТЙШСаЪБЯТНЕЕНдМ3КСФІЖћ/Щ§ЃЌДгЖјжЄУїСЫNNC2215ЖдПЙЕЭбЊЬЧЕФБЃЛЄзїгУЁЃ
вЉДњЖЏСІбЇ/вЉаЇбЇФЃаЭ
ЭЈЙ§ЖджэЕФвЉаЇбЇЪ§ОнгІгУзюаЁФЃаЭНјааЗжЮіЃЌИУФЃаЭвдЖЈСПЗНЪНУшЪіСЫЦЯЬбЬЧЖЏСІбЇКЭвШЕКЫиЛюадЃЈВЙГфаХЯЂКЭРЉеЙЪ§ОнЭМ5aЃЉЁЃНсКЯвЉДњЖЏСІбЇФЃаЭЃЈВЙГфаХЯЂКЭРЉеЙЪ§ОнБэ1bЃЉЃЌзюаЁФЃаЭгУгкСПЛЏNNC2215ЫцбЊНЌЦЯЬбЬЧБфЛЏЕФвШЕКЫиЛюадБфЛЏЁЃЭЈЙ§ЖдвЛжЛНгЪм1.72ЦЄФІЖћ/ЧЇПЫ/ЗжжгNNC2215МССПЕФLYDжэЕФЦЯЬбЬЧЪ§ОнНјааФтКЯЃЌЕУЕНСЫзюМбФтКЯЃЈЭМ4dЃЉЁЃЖдгкетжЛжэЃЌЕБЭЃжЙЦЯЬбЬЧЪфзЂЪБЃЌбЊНЌЦЯЬбЬЧДг8.5КСФІЖћ/Щ§НЕжС3.8КСФІЖћ/Щ§ЃЌSIжИЪ§Дг140%НЕжС70%ЃЌБэУїдкЯрЙиЦЯЬбЬЧЗЖЮЇФкNNC2215МЄЛюЕФЯджјЖЏЬЌЗЖЮЇЁЃЖдгквШЕКЫиЕТЙШСаЃЌSIжИЪ§ЪМжеЮЊ100%ЃЌШчЭМ4dжаЕФЫЎЦНащЯпЫљЪОЁЃФЃаЭБэУїЃЌдк3жС15КСФІЖћ/Щ§ЦЯЬбЬЧЗЖЮЇФкЃЌбЊНЌЦЯЬбЬЧгыSIжИЪ§жЎМфЕФЙиЯЕЪЧЯпадЕФЃЌаБТЪЮЊУПКСФІЖћ/Щ§ЦЯЬбЬЧ14.6%ЃЈРЉеЙЪ§ОнЭМ5bЃЉЁЃ
дкSTZ-ЬЧФђВЁДѓЪѓжаЕФЦЯЬбЬЧФЭЪмад
вШЕКЫиШБЗІЕФСДыхзєОњЫиЃЈSTZЃЉЬЧФђВЁДѓЪѓБЛгУРДдкВЛИЩШХФкдДадвШЕКЫиЪЭЗХЕФЧщПіЯТЃЌЭЈЙ§ОВТіGTTбаОПЦЯЬбЬЧгеЕМЕФNNC2215МЄЛюЁЃИєвЙНћЪГЕФДѓЪѓЭЈЙ§ОВТівдШ§жжВЛЭЌЕФКуЖЈФІЖћЫйТЪЪфзЂNNC2215ЛђШЫвШЕКЫиЃЌвдНЋбЊНЌЦЯЬбЬЧДгЁн15КСФІЖћ/Щ§НЕжСдЄЖЈФПБъ5.7КСФІЖћ/Щ§ЁЃЕБЦЯЬбЬЧЪфзЂЫйТЪЮШЖЈЪБЃЌНјааGTTЃЌЭЈЙ§дкИіЬхЮШЖЈзДЬЌЦЯЬбЬЧЪфзЂЫйТЪЕФЛљДЁЩЯЖюЭтЪфзЂ25КСПЫ/ЧЇПЫ/ЗжжгЕФЦЯЬбЬЧ60ЗжжгЃЈЭМ5aЃЉЃЌВЂМЧТМНсЙћЁЃ
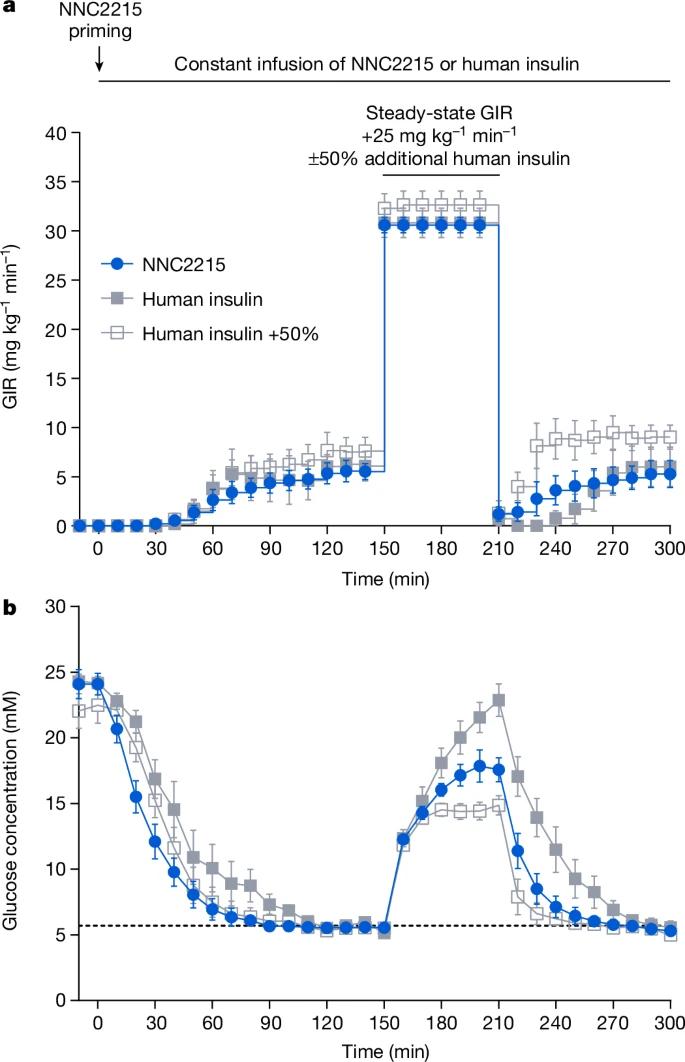
ЭМ5ЃКСДыхзєОњЫиЬЧФђВЁДѓЪѓжаNNC2215дкЦЯЬбЬЧФЭСПВтЪдЃЈGTTЃЉЦкМфЕФЦЯЬбЬЧгеЕММЄЛюЁЃ
a,bЃЌдкНгЪмОВТіЪфзЂNNC2215ЃЈ84 pmol kg⁻¹ min⁻¹ЃЉЁЂШЫвШЕКЫиЃЈ20 pmol kg⁻¹ min⁻¹ЃЉЛђШЫвШЕКЫиЃЈ20 pmol kg⁻¹ min⁻¹ЃЉМгЩЯGTTЦкМфЖюЭт10 pmol kg⁻¹ min⁻¹ЃЈ+50%ЃЉЕФСДыхзєОњЫиЬЧФђВЁДѓЪѓжаЃЌGTTЧАЃЈ0ЈC150ЗжжгЃЉЁЂGTTЦкМфЃЈ150ЈC210ЗжжгЃЉКЭGTTКѓЃЈ210ЈC300ЗжжгЃЉЕФЦЯЬбЬЧЪфзЂТЪЃЈGIRЃЉЃЈaЃЉКЭбЊНЌЦЯЬбЬЧЃЈbЃЉЧњЯпЁЃЪ§ОнБэЪОЮЊЦНОљжЕЁРБъзМЮѓЃЌn=5ЃЈNNC2215КЭШЫвШЕКЫиЃЉКЭn=7ЃЈШЫвШЕКЫи+50%ЃЉДѓЪѓЁЃ
ЭМ5ЯдЪОСЫдкGTTжЎЧАОпгаЯрЫЦЮШЬЌЦЯЬбЬЧЪфзЂТЪЃЈМДЯрЫЦЕФвШЕКЫиаЇгІЃЉЕФSTZЬЧФђВЁДѓЪѓЕФЪ§ОнзгМЏЁЃШчЭМ5bЫљЪОЃЌдкGTTЦкМфЃЌгыШЫРрвШЕКЫизщЯрБШЃЌNNC2215зщЕФбЊНЌЦЯЬбЬЧдіМгДѓдММѕЩйСЫ20%ЃЈЦНОљжЕЁРБъзМЮѓЃЌ18.3ЁР1.1ЖдБШ22.9ЁР1.3 mMЃЌP<0.02ЃЉЃЌБэУїдкИпЦЯЬбЬЧХЈЖШЯТNNC2215ЕФвШЕКЫиаЇгІИќДѓЃЌЖјвШЕКЫиБЉТЖСПУЛгаБфЛЏЁЃЮЊСЫСПЛЏ50%ЖюЭтШЫРрвШЕКЫидкGTTЦкМфФмМѕЩйЖрЩйзюДѓбЊНЌЦЯЬбЬЧХЈЖШЃЌСэвЛзщДѓЪѓдкЦфКуЖЈЪфзЂТЪЕФЛљДЁЩЯИјгшСЫ50%ЕФЖюЭтШЫРрвШЕКЫиЃЌМДдкGTTЦкМфЮЊ20+10 pmol kg⁻¹ min⁻¹ЁЃгыЕШФІЖћЪфзЂТЪЕФШЫРрвШЕКЫиЯрБШЃЌ50%ЖюЭтШЫРрвШЕКЫиЕМжТЕФзюДѓбЊНЌЦЯЬбЬЧХЈЖШЯджјНЕЕЭЃЈдМ34%ЃЉЃЈЦНОљжЕЁРБъзМЮѓЃЌ15.1ЁР0.7ЖдБШ22.9ЁР1.3 mMЃЌP<0.0002ЃЉЃЌВЂЧвгыNNC2215ЯрБШЃЌзюДѓбЊНЌЦЯЬбЬЧвВЧуЯђгкНјвЛВННЕЕЭЃЈЦНОљжЕЁРБъзМЮѓЃЌ15.1ЁР0.7ЖдБШ18.3ЁР1.1 mMЃЌP=0.08ЃЉЃЈЭМ5bЃЉЁЃ
ЖдЭъећЪ§ОнМЏЕФЗжЮіЯдЪОЃЌдкВЛЭЌвШЕКЫиМССПВтЪджаЃЌгЩNNC2215КЭ50%ЖюЭтШЫРрвШЕКЫив§Ц№ЕФзюДѓбЊНЌЦЯЬбЬЧХЈЖШНЕЕЭгыШЫвШЕКЫидкЕШФІЖћЪфзЂТЪЯТЕФНЕЕЭЯрЕБЁЃдкGTTЦкМфЃЌNNC2215ЕФЦЯЬбЬЧгеЕМдіМгЕФвШЕКЫиЛюадЙРМЦЯрЕБгкШЫРрвШЕКЫиМССПдМ30%ЕФдіМгЃЈРЉеЙЪ§ОнЭМ6ЃЉЁЃ
ЬжТл
вШЕКЫиХфБИСЫвЛИіЖдЦЯЬбЬЧУєИаЕФПЊЙиЃЌИУПЊЙигЩгыLysB29НсКЯЕФДѓЛЗКЭЭЈЙ§ЖЬСДгыB1PheНсКЯЕФO1-ЬЧмезщГЩЁЃЪЙгУжЪЦзЗжЮіЦРЙРСЫNNC2215гыЦЯЬбЬЧЕФНсКЯЃЌНсЙћЯдЪОНсКЯЧњЯпзюЖИЕФВПЗжгыЕЭбЊЬЧЗЖЮЇЯрЦЅХфЃЌМДЕЭгк4 mMЦЯЬбЬЧЁЃВтСПСЫдк0ЕН20 mMЦЯЬбЬЧХЈЖШЗЖЮЇФкЃЌNNC2215ЖдвШЕКЫиЪмЬхЕФЦЯЬбЬЧУєИаНсКЯЃЌЗЂЯжNNC2215ЕФЦЯЬбЬЧЗДгІадБШжЎЧАГЂЪдЮЊвШЕКЫиХфБИЦЯЬбЬЧУєИаПЊЙиЕФБЈИцвЊЧПЕУЖрЁЃЭЈЙ§3DЗжзгНЈФЃбаОППЩвдКЯРэНтЪЭNNC2215гывШЕКЫиЪмЬхЯрЛЅзїгУЕФЦЯЬбЬЧДйНјЕїНкЃЌПЩЪгЛЏеЙЪОСЫДІгкЙиБезДЬЌЕФNNC2215ШчКЮИЩШХгывШЕКЫиЪмЬхІС-CTгђЕФНсКЯЁЃЮЊСЫзїЮЊвЉЮяЪЙгУЃЌетИіПЊЙигІИУЖдЬЧФђВЁЛМепжаГіЯжЕФЦЯЬбЬЧХЈЖШЃЈМДдМ2ЕН20ЈC30 mMЃЉзіГіЯьгІЁЃДѓЛЗЕФЦЯЬбЬЧЬивьаджЎЧАвбОбаОПЙ§ЃЌЕЋВЛФмХХГ§ЫќПЩФмЛсгыЬЧЛљЛЏЕААзЗЂЩњвЛаЉНсКЯЁЃШЛЖјЃЌЕБЙлВьвШЕКЫиЪмЬхЪБЃЌЫќЪЧвЛжжЬЧЛљЛЏЕААзЃЌВЂЧвЪЧЪЙгУКЌгаТѓХпФ§МЏЫиЃЈWGAЃЉЕФЬЧЛљЛЏНсКЯжљНјааАыДПЛЏЕФЃЌШчЙћNNC2215гывШЕКЫиЪмЬхЕФОлЬЧНсКЯЃЌФЧУДНіКЌДѓЛЗЕФЖдееЛЏКЯЮяNNC2215aгІИУдкНсКЯВтЖЈжаБэЯжГіЦЯЬбЬЧУєИаадЃЌЖјЪЕМЪЩЯВЂУЛгаЁЃетБэУїNNC2215дкКмДѓГЬЖШЩЯВЛЛсгыЬЧЛљЛЏЕААзНсКЯЁЃ
дквЛИіМђЕЅЕФМБадДѓЪѓФЃаЭжажЇГжСЫЬхФкЕФЦЯЬбЬЧУєИаадЃЌЪЙгУl-ЦЯЬбЬЧМССПРДДЅЗЂNNC2215ЕФвШЕКЫиаЇгІЖјВЛДЬМЄФкдДадвШЕКЫиЪЭЗХЁЃЭЈЙ§l-ЦЯЬбЬЧМЄЛюЕФNNC2215ЕМжТd-ЦЯЬбЬЧНЕЕЭКЭNNC2215ЧхГ§ГЪМССПвРРЕадЁЃетвЛЗНАИЪмЕНЪЙгУІС-МзЛљ-d-ИЪТЖЬЧДЅЗЂвРРЕгкЯШЧАУшЪіЕФИЪТЖЬЧЪмЬхдРэЕФЦЯЬбЬЧУєИавШЕКЫибмЩњЮяЕФЦєЗЂЁЃ
жЕЕУзЂвтЕФЪЧЃЌNNC2215ЕФЦЯЬбЬЧУєИавШЕКЫиЪмЬхНсКЯКЭЯИАћаЇгІзЊЛЏЮЊдкгЩЩњГЄвжЫиКЭвШИпбЊЬЧЫиЬцДњЪфзЂгеЕМЕФМБадЬЧФђВЁбљжэФЃаЭжаЙлВьЕНЕФЬхФкЕЭбЊЬЧБЃЛЄаЇгІЁЃЖдгкЭЌбљЕФНЕЬЧаЇЙћПЊЪМЪБЃЌЕБd-ЦЯЬбЬЧЪфзЂЙиБеЪБЃЌЮвУЧЙлВьЕНбЊНЌЦЯЬбЬЧЯТНЕЃЌЕЋNNC2215ЕФЯТНЕЗљЖШБШвШЕКЫиdegludecаЁЃЌзюЕЭЦЯЬбЬЧжЕЦНОљдМЮЊ4.5 mMЃЌЖјвШЕКЫиdegludecЕЭгк3 mMЁЃдкЫљгаВтЪдЕФМССПЫЎЦНЩЯЙлВьЕНNNC2215гывШЕКЫиdegludecжЎМфЦЯЬбЬЧЯТНЕЕФВювьЃЌЙРМЦдкжиЕўЕФЦ№ЪМЦЯЬбЬЧЗЖЮЇФкдМЮЊ1.8 mMЃЈРЉеЙЪ§ОнЭМ3ЃЉЁЃЕБвШЕКЫиМССПгыЪЕМЪашЧѓВЛЦЅХфЪБЃЌетжждкЕЭбЊЬЧЫЎЦНЪБМѕЩйВЛРэЯыЕФбЊНЌЦЯЬбЬЧНЕЕЭаЇгІЕФГЬЖШдЄМЦЛсИјЯёNNC2215етбљЕФвШЕКЫиХМСЊЮяДјРДЯджјгХЪЦЃЌМѕЩйвШЕКЫижЮСЦЙЬгаЕФЕЭбЊЬЧЗчЯеЁЃ
ЮвУЧЛЙЙлВьЕНдкSTZЬЧФђВЁДѓЪѓжаЕФЦЯЬбЬЧЬєеНЦкМфЃЌNNC2215ЕФМЄЛюЯрЕБгкЖюЭт30%ЕФШЫвШЕКЫиаЇЙћЁЃетжжгЩЦЯЬбЬЧгеЕМЕФNNC2215МЄЛюБэУїЫќПЩФмгажњгкМѕЩйВЭКѓЕФвЛаЉЦЯЬбЬЧВЈЖЏЁЃвђДЫЃЌПЩФмашвЊНЯЩйЕФПьЫйзїгУвШЕКЫиМССПРДгІЖдНЯДѓЕФВЭЪГЁЃНЯаЁМССПЕФВЭЪБвШЕКЫигыNNC2215НсКЯЪЙгУПЩФмЧБдкЕиМѕЩйгЩПьЫйзїгУвШЕКЫив§Ц№ЕФЕЭбЊЬЧЗчЯеЃЌВЂзмЬхЩЯдЪаэИќНєУмЕФЦЯЬбЬЧПижЦЖјЮоашЕЃаФЕЭбЊЬЧЁЃ
дкжэжаЖдвЉДњЖЏСІбЇ/вЉаЇбЇЪ§ОнЕФНЈФЃСПЛЏСЫNNC2215дкЬхФкЕФЖЏЬЌЗЖЮЇЁЃИљОнвЉДњЖЏСІбЇ/вЉаЇбЇНЈФЃЃЌдкгыЬЧФђВЁЯрЙиЕФЦЯЬбЬЧЗЖЮЇФкЃЌвШЕКЫиЛюаджИЪ§дк3 mMЦЯЬбЬЧЪБдМЮЊ60%ЃЌдк20 mMЦЯЬбЬЧЪБЮЊ290%ЃЈРЉеЙЪ§ОнЭМ5bЃЉЃЌМД5БЖЕФЗЖЮЇЃЌетгыЬхЭтЗЂЯжЕФ3КЭ20 mMЦЯЬбЬЧжЎМфЕФвШЕКЫиЪмЬхЧзКЭСІ3.2БЖВювьвЛжТЁЃNNC2215дкЬхЭтКЭЬхФкЦЯЬбЬЧУєИаадЕФетжжвЛжТаджЇГжСЫОпгаРрЫЦNNC2215ЬиадЕФЛЏКЯЮяПЩФмзЊЛЏЮЊШЫРрЪЙгУЕФЧБСІЁЃ
змжЎЃЌЯёNNC2215етбљЕФвШЕКЫиХМСЊЮяЭЈЙ§ЧБдкЕиНЕЕЭЕЭбЊЬЧЗчЯеКЭВПЗжТњзуВЭЪБПьЫйзїгУвШЕКЫиЕФашЧѓЃЌгаЭћИФЩЦЬЧФђВЁЕФжЮСЦЁЃетСНИіЬиадЕФНсКЯгІИУдЪаэгыФПЧАЕФвШЕКЫиСЦЗЈЯрБШНјааИќЛ§МЋЕФвШЕКЫиЕЮЖЈЃЌвдЪЕЯже§ГЃЕФЦЯЬбЬЧЫЎЦНЖјВЛдіМгЕЭбЊЬЧЕФЗчЯеЁЃетПЩФмЛсИФЩЦгыЬЧФђВЁЯрЙиЕФЖЬЦкКЭГЄЦкЗчЯеМАВЂЗЂжЂЁЃзмЕФРДЫЕЃЌNNC2215еЙЪОСЫШчКЮЩшМЦЗжзгПЊЙивдЪЙЗжзгЩњЮяЛюадФмЙЛзджїПижЦдкСэвЛИіЗжзгБфЛЏХЈЖШЯТЕФЯьгІЃЌМДЪЙЪЧЯёбЊЬЧЫЎЦНетбљЯСеЕФЗЖЮЇЁЃ
https://wap.sciencenet.cn/blog-41174-1456161.html
ЩЯвЛЦЊЃКвћЪГЕїНкУтвпЙІФмЕФдвђ
ЯТвЛЦЊЃКЮвЙњЯждкгЕгаЪРНчЩЯзюЧПДѓЕФЕчзшДХЬњЁЖздШЛЁЗ