
 精选
精选
【内容提要】Kay等人探讨了园艺学在植物保护中的作用,特别是植物园在繁殖和保护珍稀植物方面的优势。蒙哥马利植物中心(MBC)苏铁植物繁殖计划重点研究了小苏铁(Microcycas calocoma)的繁殖和定植技术。该种因市场需求旺盛而面临濒危,MBC通过人工繁殖和推广,有助于支持其自然种群的就地保护。作者还讨论了植物园与产业界合作的新模式,以及市场力量如何影响珍稀植物的供应和保护。通过拍卖种子,小苏铁种子价格下降,供应量增加,市场得到满足,预计未来小苏铁的采集将不再具有经济吸引力。我国国家植物园应利用植物园的专业知识技术与基础设施(如温室、苗圃和种子库),开展物种保护。其保护园艺体系涵盖植物迁地保护全流程的科学技术体系,包括植物引种采样策略制定、栽培繁殖技术开发、种质保存、移栽与定植规范管理,以及多站点微生境适应性管理研究与技术应用等工作范畴,探索受威胁植物综合保护并满足市场需求策略。

植物园正日益致力于推动植物保护工作(Oldfield, 2010 等),并且与动物园类似,许多植物园已将保护确立为其核心使命之一。保护园艺学(Conservation horticulture)是利用植物园员工专业知识与现有基础设施(Kramer et al., 2011)的工作领域。这也是传统上并非由其他组织承担的保护任务(Griffith et al., 2011)。成熟的迁地保护园艺技术(ex situ conservation horticulture),使该领域成为现代公共植物园明确的发展方向。
苏铁类植物(Cycadales)普遍受到栖息地丧失的威胁。对苏铁持续存在的另一威胁是野外非法采集(Donaldson, 2003)。苏铁以其奇特的形态、古老性以及与恐龙的联系吸引着公众。它们在植物进化、形态学及保护议题的研究与教育中也具有重要价值。因此,活体苏铁是公共植物园和温室的重点展示植物。
所有苏铁均为雌雄异株且依赖昆虫传粉。由于传粉者通常不存在于人工环境,这需要植物园工作人员进行主动的人工繁殖。因此,苏铁类植物包含了一些保护园艺学最典型的案例。最著名的是伍德氏非洲铁(Encephalartos woodii),该物种目前仅存于栽培环境中,所有植株均为克隆繁殖的雄性个体,源自二十世纪初被迁入保护性栽培的最后一株野生植株(Prakash et al., 2008)。其他案例还包括德保苏铁(Cycas debaoensis)(Tang et al., 2004)、双子苏铁(Dioon edule)(Vovides, 2010)、泽米铁属植物(Zamia spp. )(Calonje et al., 2010; Griffith et al., 2010)以及密克罗尼西亚苏铁(Cycas micronesica)(Griffith and Calonje, 2007)的繁殖项目。为进一步推动苏铁保护园艺学的发展,蒙哥马利植物中心(MBC)种子库将工作重点放在苏铁的繁殖与分发上。MBC生产的苏铁种子与公共植物园(见表1)、学校、保护主义者和研究人员共享。在完成这些分发之后,收藏家和园艺产业才有机会合法购买这些稀有物种的种子。来自这些购买的数据有助于了解该物种在栽培环境中的供需关系,这对防止对野生种群的过度采集具有指导意义。

1 模型系统:小苏铁
小苏铁(Palma corcho)是小苏铁属的唯一物种,为古巴特有苏铁,与泽米铁属(Zamia)近缘,仅分布于比那尔德里奥省。该树状苏铁植物可高达 11 米(Lazcano, 2007),叶片呈醒目的截形,小叶片反折、半下垂,使整株植物形态规整,近乎修剪造型。它是新世界备受追捧的珍贵观赏苏铁(Jones, 1993),深受收藏家青睐。
原生种群被评定为“极危”(critically endangered),这是世界自然保护联盟对野生物种设定的最高风险等级,原因是近期种群数量急剧下降(Bösenberg, 2009;Peña et al., 1998)。现存野生植株的估算数量从 600 株(Jones, 1993;Whitelock, 2002)到 1 540 株(Bösenberg, 2009)不等,甚至可能高达 4 000 株,但预计仍将继续减少(Bösenberg, 2009)。专家指出,导致衰退的因素包括:作为观赏植物的非法采集、生境破坏、自然繁殖率下降、传粉者减少以及外来食草动物的掠食(Lazcano-Lara, 2004;Pinares et al., 2009;Vovides et al., 1997)。正如野生 Palma corcho 权威学者 Julio Lazcano(2007)所言,该物种的保护策略之一是“通过迁地栽培建立种质资源,以备未来再引入、交流或受控销售”。
为切实推进这一保护工作,本文以 Palma corcho 为范例,报告了一项植物园苏铁繁育项目的案例研究。该模式系统聚焦于一种极度珍稀、备受追捧且繁殖条件严苛的植物,其可繁殖的种源极为有限。
2 MBC 小苏铁种源信息
1915 年,实业家詹姆斯·迪林(James Deering)为装点其位于佛罗里达州迈阿密的庄园——维斯卡亚(Vizcaya),购进一大批成龄王棕(Roystonea)。这些王棕由古巴的 H·A·范·赫尔曼(H. A. van Hermann)提供(Gayer,无日期)。范·赫尔曼时任古巴国家园艺学会(CNHS)财务主管,并兼任古巴农业部官员(CNHS,1911)。1914 年末,他在野外采集了小苏铁(palma corcho)(图 1)。运抵维斯卡亚的这批苗木中,夹杂了几株小苏铁,当时干高 3–5 英尺(Sturrock,1915),估计树龄约 40 年,系野生挖掘(Calonje,2007)。苏铁被栽入大缸,陈列在迪林宅邸正门及赤陶花园中。
1932 年,罗伯特·蒙哥马利上校(Colonel Robert Montgomery)开始创建椰林棕榈园(后改名蒙哥马利植物中心,MBC)。为此,他以 90 美元(约合 2010 年的 1 450 美元)从迪林家族购入两株小苏铁(Calonje,2007)。当时两株干高分别为 5 英尺和 8 英尺。较小的那株(登录号 RM384)至今仍在 MBC 生长(图 2、图 3)。这株雄株(昵称“祖父”)是花粉的重要来源。2010 年,它被佛罗里达州林业厅命名为“冠军树”,可能是古巴境外最大的小苏铁。蒙哥马利上校在 1938 年创办费柴尔德热带植物园(Fairchild Tropical Botanic Garden, FTBG)时,将较大的一株(同为雄株)捐赠给了 FTBG。

图 1 所示小苏铁标本(van Hermann 564,藏于哈瓦那古巴生态与系统学研究所植物标本室)为 1914 年范·赫尔曼在古巴比那尔德里奥野外采集的雄株,摄于数月后运往维斯卡亚之前。
由于当时美国境内所有已知小苏铁均为雄株,1959 年FTBG的斯坦利·基姆(Stanley Kiem)与古巴园艺家马西亚尔·特鲁芬(Marcial Truffin)专程赴古巴采集材料用于繁殖(Kiem,1963)。最终仅两株存活并开花:一株雄株于 1974 年首次抽球穗,另一株雌株于 1977 年抽球穗。1977 年二者授粉成功,首次收获种子(Calonje,2007)。1991 年,这株唯一的雌株遭雷击死亡。1994 年,由 1977 年种子育成的植株开始抽雌球穗。自 1995 年起,MBC 对这些雌株进行人工授粉,并于 1996 年首次对外分发种子。因此,过去二十年的全部种子皆源自这批 1977 年园内繁育的雌株。栽培材料的匮乏在植物园保护国际(BGCI, 2011)的数据中亦可见一斑:全球仅 32 家植物园保存小苏铁。

图 2:1932 年拍摄的照片中,登录号 RM384 的小苏铁作为首批苏铁之一,栽植于佛罗里达州科勒尔盖布尔斯的蒙哥马利植物中心。来自迈尔斯堡的著名园艺师 J·E·亨德利(J. E. Hendry Jr.)作比例尺。

图 3:2010 年照片显示,这株登录号 RM384 的小苏铁冠幅已达 4 米(13.1 英尺),估计树龄约 135 岁。费柴尔德热带植物园退休园长J. Popenoe 与植株。
迄今,MBC 已向 13 家公共园艺机构提供了小苏铁材料(见表 1),这 13 家与 BGCI 统计的 32 家并不完全重叠。
这段历史凸显了冗余规划、植株交换及积极繁育活植物收藏在缓解自然灾害意外损失中的重要作用(Bergquist, 2009, 2010;Dosmann & Del Tredici, 2003)。简言之,MBC 作为繁殖母本的小苏铁活植物收藏来源多元,其建立并非以遗传多样性或再引入为最初目的(Maunder et al., 2004;Radford et al., 2003)。
3 物候与授粉
在原生地,雄球花与雌球花的发育需 4 个月:9 月散粉/雌球花可受粉,种子于翌年 7 月成熟,历时 10 个月(Whitelock, 2002)。栽培条件下,据 Norstog 与 Nicholls(2002)报道,雌株隔年单抽一球穗,9 月受粉,球花持续至次年 6 月;雄球花 7–8 月成熟,9 月上旬散粉。我们在迈阿密观察到,小苏铁雌球花的可受粉高峰在 8 月,7 月与 9 月亦偶有发生(图 4)。部分雌株可连年抽球穗,一次偶见 1–2 枚雌球花,通常由不同顶芽抽出。MBC 收藏的雄球花于 8–9 月散粉。Whitelock(2002)称 8 年生植株即可抽球穗,Norstog 与 Nicholls(2002)则认为需 10–15 年;MBC 记录中,最早雌球穗出现于播后 18 年,最早雄球穗为 9 年。
苏铁球花在可受粉期常显著升温(热发生)(Tang et al., 1987),并伴随挥发性气味化合物(Tang, 1993)。这些气味为吸引昆虫(蓟马 Thysanoptera、象甲/甲虫 Coleoptera)设计,但人亦可闻到,常被描述为“霉味、果香或甜香”(Whitelock, 2002)。澳洲亮叶大泽米铁(Macrozamia lucida)通过气味吸引并驱离传粉者以助花粉转移(Terry et al., 2007)。我们观察到小苏铁雌雄球花在散粉时均散发霉味;Whitelock(2002)形容此味“强烈难闻”。MBC 的成熟雌球花因成熟假种皮而散发部分人认为刺鼻的气味(个人观察)。小苏铁的传粉者埃斯佩兰萨象甲(Pharaxonotha esperanzae)直至 2005 年才被正式记录描述(Chavez & Genaro, 2005),此前曾被认为近乎灭绝(Vovides et al., 1997)。
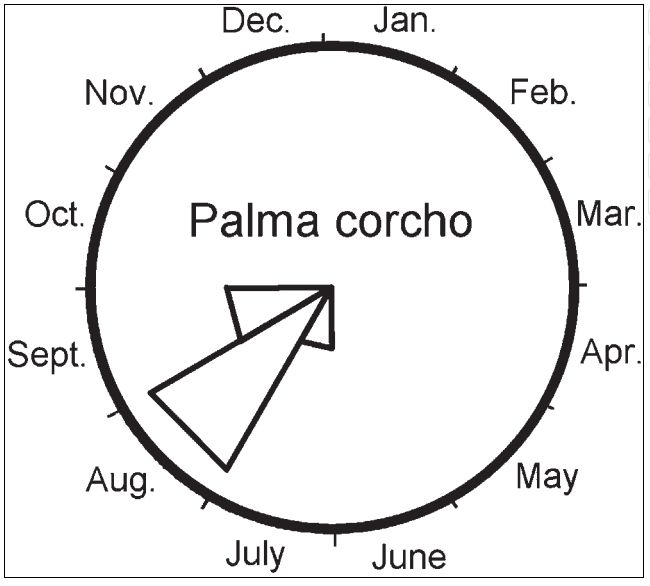
图 4. 小苏铁生殖物候图(改绘自 Griffith et al., 2011b)。扇形表示 13 年间 21 株 73 次生殖事件中,各月雌球花可受粉或雄球花散粉的比例。观察总次数 156 次。生殖事件始于 7 月,8 月达峰,9 月仍可见。
4 雄性球果——花粉采集、储存与活性检测
花粉采集: MBC 活体植物收藏中的雄性球果主要在八月成熟并释放花粉。在此过程中,球果伸长,小孢子叶分离,花粉囊(小孢子囊)裂开释放花粉。若此时雌性球果处于可授粉状态,则直接使用新鲜采集的花粉进行授粉;若雌性球果尚未可授粉,则采集花粉并进行处理以备储存。
当雄性球果开始释放花粉时,将其从植株上取下,置于铺有蜡质屠夫纸的托盘上(图 5),放置于阴凉处。必须与其他球果隔离放置,并避免通风。工作人员每日用小型木槌敲打球果约 4 天,以促进花粉释放。
储存: 将收集到的花粉清除杂质,装入小纸袋中,随后放入装有硅胶干燥剂的密封玻璃罐内。将玻璃罐置于冰箱中至少 2 天,以吸收花粉中的水分,之后可转移至 –2 ℃ 的冷冻柜中储存。在此条件下,花粉活性最长可保持 3 年。推荐使用非自动除霜的家用冰柜,因其温度更恒定。不过,若花粉经妥善处理并置于密封罐中,在自动除霜冰柜中也能保存良好(Hanna, 1994)。MBC 定期将花粉捐赠给其他植物园(如佛罗里达州马纳拉潘市的 FTBG 和 双子花园(Gemini Garden))以及苗圃。

图 5. 这株小苏铁的雄性球果正在释放花粉。花粉将被收集、干燥并在冷冻温度下储存以备将来使用。
活性检测:花粉活性可通过制备苯胺蓝染色玻片进行检测。静置 12–24 小时后,在复式显微镜下观察,具有活性的花粉粒将呈现深蓝色(图 6)。检测采用苯酚/乳酸中苯胺蓝染色的标准化方案(Hauser and Morrison, 1964)。通常会对储存超过 2 年的花粉进行活性检测,活性低(通常低于 25%)的花粉将被弃用。
5 雌性球果——可授粉期与授粉
当雌性球果进入可授粉期时,其大孢子叶之间会出现显著的纵向裂缝。雌性球果的可授粉期通常至少持续一周。尽管许多苏铁采用湿式授粉(wet pollination)法繁殖,但我们对小苏铁(palma corcho)采用干式授粉(dry pollination)法,因其胚胎已被观察到易受一种真菌病原体侵染(个人观察)。该病原体目前正在鉴定中。
在可授粉期内,工作人员会分三个不同的日期,使用手持球囊式吹粉器(hand-bulb compression tool)将花粉吹入裂缝中(图 7)。随着种子成熟,球果会变得下垂且沉重,因此我们常会使用木质支架加以支撑(图 7),以防其因重量而提前脱落。
6 种子收集、活性检测、萌发与养护
雌性球果平均需要 310 天才能成熟。球果成熟过程中,我们尽量让种子留在球果上,直至其自然脱落(图 8)。种子收集通常持续 8 天。每个球果可产生 500 至 600 粒种子。
种子活性通过剖切约 20 粒种子并观察是否存在胚胎来确定。并非所有含胚胎的种子都能萌发,但由于种子活性期短,在分发前进行萌发测试并不可行,因此活性检测是判断种子是否具有萌发潜力的最佳可用指标。过去的种子活性率在 0% 到 90% 之间波动,平均约为 45%。
小苏铁种子具有高度顽拗性(recalcitrant),必须在非常新鲜时播种,最迟不超过 4 个月(Whitelock, 2002)。在适宜条件下,种子 2–3 周内即可萌发。种子分发时通常附带肉质种皮(sarcotesta)(即未清洗状态),但最佳做法是由接收方立即清洗并播种。
在 MBC,我们(在清洗后)使用过氧化氢(Hydrogen peroxide,商品名 ZeroTol;BioSafe Systems 公司,康涅狄格州东哈特福德市)处理种子以防治真菌。萌发需要排水良好的介质,我们使用硅砂进行萌发,后续换盆时则使用无机介质(如:煅烧粘土颗粒 Turface MVP;Turface Athletics 公司,伊利诺伊州布法罗格罗夫市)(Calonje et al., 2010),以维持长期排水性并减少有机介质可能引发的腐烂问题。
萌发率(始终低于活性率)因栽培实践差异而不同,在 MBC 的萌发率范围在 0% 到 60% 之间,平均为 26%。种子清洗后直接播种于混播盆(community pots)中,首次萌发发生在 5 个月内。MBC 初期采用混播盆(一个 2 加仑容器中播种 10–20 粒种子),因为经验表明此法腐烂问题较少(相较于单粒种子单容器播种)。这一优势很可能源于混播盆可使用更深的容器。幼苗在萌发后 6 至 8 个月从混播盆移栽至独立盆中。
种子和幼苗使用城市自来水进行人工浇水,每周三次(或视需要而定)。苗圃幼苗在每次换盆时(平均每 6 个月一次)施用 18N–2.6P–6.6K 配方的缓释肥料(商品名 Nutricote;Florikan 公司,佛罗里达州萨拉索塔市)。在 MBC,萌发工作在 20 × 60 英尺(约 6×18米) 的玻璃温室中进行,温度维持在 60–80°F(15–27℃)。
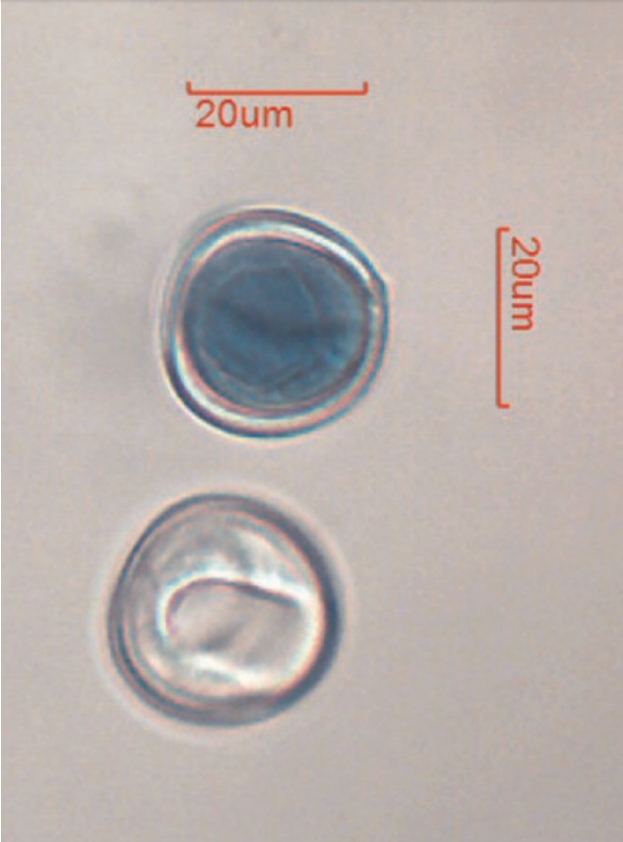
图 6. 储存的小苏铁花粉定期进行活性检测。经苯胺蓝染色(Hauser and Morrison, 1964)后,活性花粉粒呈蓝色(图中上方颗粒)。此花粉(引种编号 77404*P)在检测前已在冷藏条件下储存 10 个月。

图 7. 蒙哥马利植物中心(佛罗里达州科勒尔盖布尔斯市)志愿者 Larry Krauss 正在为可授粉期的雌性球果(引种编号 77404*G,1977 年实生苗之一)授粉。注意球果鳞片间的垂直裂缝,这表明其处于可授粉期。

图 8. 图中小苏铁(引种编号 77404*G)的雌性球果(摄于六月下旬)已结出成熟种子。球果鳞片自然脱落释放成熟种子。蓝色胶带(右下角,叶丛后方)标记表明此球果已接受人工授粉。
据报道,小苏铁植株在萌发后的头 2 年内极易感染真菌。MBC 尚未观察到此问题,这很可能归因于持续使用排水性极佳的介质,尤其是无机介质(图 9)。此阶段也使用 ZeroTol 直接喷洒植株表面以防治真菌。温室内良好的空气流通(即避免过度拥挤)也有助益。
小苏铁在苗圃中还易受咖啡盔蚧(Saissetia coffeae)侵害。对于咖啡盔蚧,先使用软毛牙刷人工清除虫体,然后施用呋虫胺(Dinotefuran,商品名 Safari;Valent Professional Products 公司,加利福尼亚州核桃溪市),这是一种广谱杀虫剂。根据制造商说明,呋虫胺也可用作灌根剂。
据称,一旦茎干成形,小苏铁是一种生长迅速的苏铁,在亚热带气候下每年可生长 10–20 厘米茎干(Whitelock, 2002)。我们在 1977 年培育的植株现高约 6 英尺(约 1.8 米)。小苏铁遇霜冻会落叶,冬季宜保持较干燥环境。
在 MBC,对有腐烂问题的小苏铁植株,会先切除受感染组织,再用过氧化氢处理。对于咖啡盔蚧,则修剪受害叶片,必要时清洗植株顶端(生长点)并施用呋虫胺。还会在树干基部周围铺设废弃咖啡渣作为覆盖物(mulch),据称此法能抑制蚧虫(Broome, 2007)。地栽植株则施用优质棕榈科专用肥料(8N–1.7P–10.0K)。
7 利用市场支持保护
1990 年代中期,蒙哥马利植物中心(MBC)与佛罗里达苗圃、种植者和景观协会(Florida Nursery, Growers and Landscape Association - FNGLA,佛罗里达州奥兰多市)建立了合作伙伴关系,旨在分发 MBC 活体植物收藏所产生的种子。丰富的、具有独特来源的繁殖材料与对新奇园艺材料的需求市场正等待着结合。通过严格遵守许可要求【《濒危野生动植物种国际贸易公约》(CITES)】和伦理规范,形成了一种创新流程:利用市场来推动保护园艺学的发展。
MBC 生产的小苏铁(palma corcho)种子多年来已分发给众多植物园(见表1)。在完成此类面向研究、教育和保护的分发之后,剩余的种子供应可以分发给国内园艺产业,以满足市场对新植株的需求。此项活动产生的收入随后可用于支持繁殖工作所需的人员和物资。
1998 年,一群园艺产业股东提供了初始资金,使 MBC 和 FNGLA 得以雇佣一名种子库协调员,作为回报,这些股东获得了当年的剩余种子。2000 年代中期启动了在线拍卖(FNGLA, 2011),使供应商和消费者都能更高效地参与市场。该拍卖也为苏铁市场提供了直接的数据来源。根据 CITES 的法律和许可框架,种子拍卖仅限于美国境内。
供应(SUPPLY):在 MBC-FNGLA 种子库成立之前,1996-98 年间 MBC 平均每年分发 560 粒 小苏铁种子(图 10)。1999 年的真菌问题导致当年未能分发。2000-02 年,MBC 能够分发种子,平均每年 950 粒。2003 年的另一场真菌问题减少了种子产量,随后在 2004 和 2005 年给予雌株休养期以进行治疗和恢复。此举效果显著,分发的种子数量缓慢上升,2007-09 年达到平均每年 1000 粒。在最近一次分发(2010 年)中达到顶峰,从三个人工授粉的球果中收获了 2155 粒种子。因此,利用活体资产(1915 年至今精心收集培育)并投资于繁殖(人员与设备),显著增加了小苏铁的供应。

图 9. 蒙哥马利植物中心(佛罗里达州科勒尔盖布尔斯市)繁殖的小苏铁幼苗种植在无机介质中。在这张 2010 年 12 月的照片中,标记为 090395 的植株是播种后 1.5 年的苗,标记为 080458 的植株是播种后 2.5 年的苗。与许多苏铁一样,小苏铁生长缓慢,幼苗期尤甚。缓慢的生长速度可能成为观赏园艺发展的障碍。
需求(DEMAND):追踪小苏铁种子的情况显示了与供应增加相对应的预期趋势:每粒种子的价格稳步下降。小苏铁种子于 2007 年首次出现在拍卖会上。种子按批次拍卖,每批数百粒不等,具体取决于每年的总量。每粒种子的平均价格首先迅速下降(图 11),从首次推出时的历史最高点,到 2008 年至 2010 年呈现持续下降的价格趋势。这仍然是 MBC 提供的所有种子中最高的价格,但供应增加的影响似乎持续压低了价格。
价值(VALUE):脱离背景孤立地看每粒种子的价格(图 11),价值下降是第一印象。但我们主张,出于保护目的,采取行动降低稀有植物的商业价值是一项积极的战略贡献。苏铁在苗圃贸易中的广泛可得性可以减少非法采集或盗窃活体收藏品的行为。对于像小苏铁这样备受觊觎的物种,压低市场价格是减少盗采的综合策略的一部分,因为它消除了(盗采的)动机。
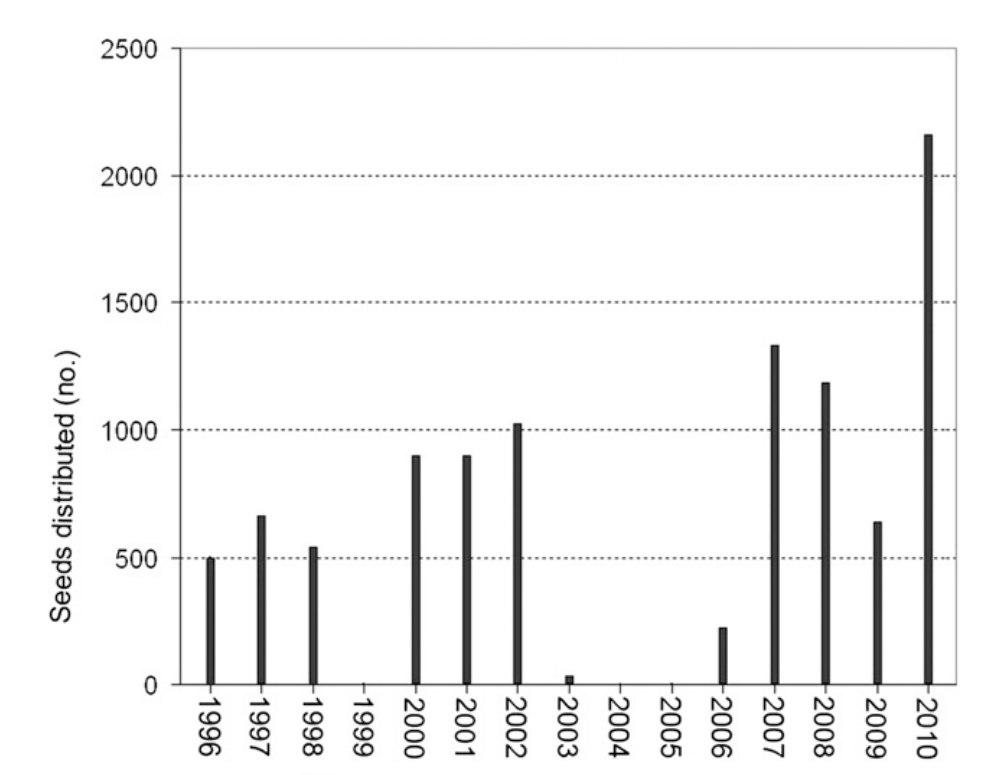
图 10. 自 1996 年以来,佛罗里达州科勒尔盖布尔斯市蒙哥马利植物中心一直分发小苏铁种子。过去 15 年,年种子产量总体呈上升趋势。1999 年及 2004-05 年因真菌病原体问题未分发种子。
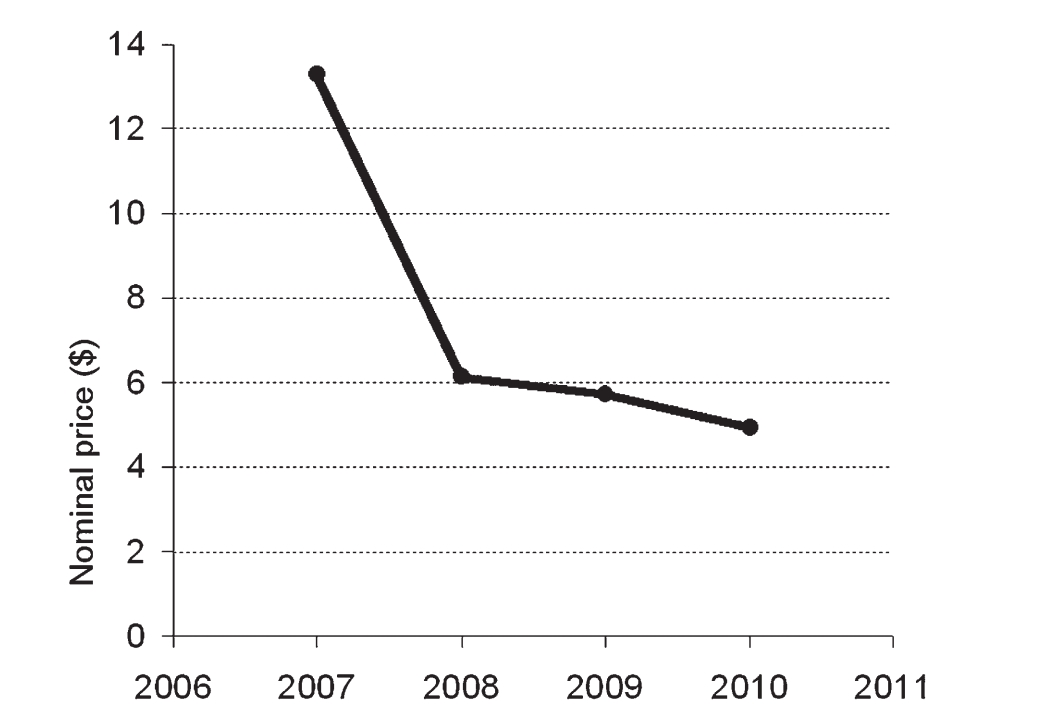
图 11. 拍卖结果显示小苏铁种子每粒的平均名义价格随时间推移而下降。即使对这些数值进行通胀调整(换算为 2007 年不变美元)或经济衰退调整(基于 2007 年不变国内生产总值),也并未改变这一趋势。2007 年首次推出的种子获得了最高的平均价格,随后价格立即大幅下跌,之后稳步下降。
预测(PROJECTION):小苏铁的未来市场会是什么样子?图 10 和图 11 的趋势显示供应量稳步增加,价格稳步下降。拍卖参与者间的非正式报告表明,通过种子库获得的种子能够萌发,其中一些已成功度过关键的幼苗阶段。因此,不仅年轻的 MBC 来源幼苗正为专业收藏家市场而培育,而且在不久的将来某个时候,越来越多的苗圃将拥有生殖成熟的植株用于其自身的繁殖活动。
根据上文报告的首个球果形成所需年龄范围(最少 8-10 年),最早在 2004-06 年就可能已有来自 MBC 种子分发的植株开始结球果,此后可能性逐年增加。随着加速苏铁生长的容器栽培技术进一步发展(例如 Calonje et al., 2010; Dehgan, 1999),更广阔的市场也可能在十年内看到足够数量且价格低廉的植株。

8 小苏铁作为模型系统
小苏铁可作为保护园艺学的一个范例,展示了如何利用和管理活体植物收藏(living collections) 以产生可衡量的成果。将其作为植物保护的模型系统能阐明某些原理,但此模型也并非适用于所有情况。下文我们将讨论此案例如何作为模型运作及其适用性。
在本案例研究中,我们探讨了一种难以获取(Jones, 1993)且面临野外过度采集威胁(Lazcano, 2007)的植物的管理。蒙哥马利植物中心(MBC)的另一项案例研究则使用了未受直接采集威胁的模型物种【匙叶棕(Leucothrinax morissii,keys thatch palm)】来探索遗传多样性的捕获(Namoff et al., 2010)。因此,当前的模型系统有潜力阐释那些采集压力构成影响因素的案例。在本研究中,我们还能看到为育种目的管理苏铁活体收藏所需的高强度资源投入(管理花粉、监测物候、主动育种和种子分发)。相比之下,匙叶棕的收藏管理则无需如此高度的专业化(Griffith and Husby, 2010)。
在当前案例中,活体收藏始于近一个世纪前,通过历史贸易、野外采集和密集繁殖缓慢发展而来。这种收藏的建立是多元化(eclectic) 的(而非系统性的),最初主要动机是展示,而非迁地保护。基于政治、法律、历史和市场的因素,尽管缺乏详细的野外来源地数据或广泛的样本量——这些是现代植物保护管理的标志(Lawrence et al., 1995)——有限的小苏铁收藏仍成为一项重要资产。促成此范例的还有其生物学特性,包括极长的世代周期和专性特化传粉(Vovides et al., 1997)。自然灾害导致的意外损失(例如 Griffith et al., 2008)在此案例中也扮演了角色;当前的小苏铁繁殖材料因1959年部分收藏的损失而进一步近交,如今仅限于1915年雄性植株和1959年植株的后代。
经济数据对此模型系统尤其具有参考价值。在所有通过在线拍卖提供的种子中,小苏铁种子在每年上市时都售价最高。这反映了该物种在收藏家心目中的珍贵地位,进而转化为苗圃对种子的需求。关于这些数据的一个潜在疑虑,可能是2008年经济危机对种子价格造成的混淆效应——2007年至2008年间的价格下跌是与供应相关,还是受整体经济状况影响?为验证此点,我们应用了校正因子,将数据转换为2007年不变美元或基于2007年不变国内生产总值。应用校正因子后,种子价格的趋势仍显示出急剧下降继而稳步贬值。此外,小苏铁拍卖在种子新鲜时进行(六月和七月),因此2008年10月的经济危机不太可能对2007-08年的种子价格下跌产生影响。
经济学与保护之间的关系已得到充分证实,主要见于动物盗猎研究(例如 Bulte and van Kooten, 1999)。对于植物,文献多集中于药用目的的过度采集,尽管瓦勒迈杉(Wollemia nobilis,wollemi pine) 的生产提供了一个非药用的范例(Trueman et al., 2007)。相关研究考察了墨西哥干旱地区(Hersch-Martínez, 1995)、喜马拉雅高山带(Chandra et al., 2006; Smith Olsen and Overgaard Larsen, 2003)以及巴西(de Melo et al., 2009; Homma, 1992)的药用植物。一个反映本研究情况的案例表明,非官方市场价格在高需求与稀缺供应下会上涨,因此增加栽培有望支持就地保护(Chandra et al., 2006)。
小苏铁模型系统能将这种保护经济学理念引入公共园艺领域。该模型表明,利用植物园历史遗留的收藏来生产和分发种子,能在很大程度上满足市场对稀有植物的需求。正如上述引用的案例所示,市场价值的降低应与野外挖掘植株动机的大幅减弱相关联。

9 局限性与改进方向
理想情况下,植物保护性收藏应致力于广泛捕获遗传多样性(Namoff et al., 2010)。鉴于现存少数植株的谱系有详细记录,蒙哥马利植物中心(MBC)小苏铁活体收藏的遗传多样性必然有限。因此,仅由这些植株产生的繁殖材料并不适合用于野外回归目的。花粉活性始终低于 100%,种子活性也始终低于 90%。考虑到已知的有限谱系(Hedrick and Kalinowski, 2000),这很可能是由于近交衰退(inbreeding depression)所致。通过引入新采集的植株来增加遗传多样性将对该系统大有裨益。
小苏铁种子接收方的反馈不尽相同。在 MBC 观察到的花粉活性、种子活性和萌发率始终分别低于 100%、90% 和 70%。种子活性已通过拍卖网站透明公开。或许由于这些种子价格高昂,投资回报的期望值也随之升高,一些误解将报告的种子活性与预期的萌发率混为一谈。此外,在某些种子中观察到真菌病原体。种子接收方表达的关切反馈有助于完善种子库项目的沟通工作,并促使 MBC 将种子样本送至佛罗里达大学的专家处进行真菌病原体分离与鉴定(目前正在进行中)。持续改进与购买方的沟通有助于阐明种子活性与萌发率之间的差异,并推广良好的栽培实践。

结论与未来展望
有效的保护园艺学必须考虑遗传多样性的捕获、数据管理、栽培和繁殖。但在本案例中,我们看到如何盘活有限的历史资产并利用市场力量,为长期的就地保护做出潜在的战略贡献。小苏铁将永远是一种美丽、独特的植物,对园艺爱好者具有特殊吸引力——但在不久的将来,任何园艺爱好者都将无法再称其为稀有或昂贵之物。
延伸阅读:Kay, J., Strader, A. A., Murphy, V., Nghiem-Phu, L., Calonje, M., & Griffith, M. P. (2011). Palma Corcho: A Case Study in Botanic Garden Conservation Horticulture and Economics. HortTechnology, 21(4), 474-483.






转载本文请联系原作者获取授权,同时请注明本文来自廖景平科学网博客。
链接地址:https://wap.sciencenet.cn/blog-38998-1495397.html?mobile=1
收藏