
 精选
精选
【内容提要】
栽培寄生植物是一项充满特殊挑战的任务,必须同时满足寄生物、宿主及其相互作用的生长条件。文章回顾了小米草属半寄生植物的栽培技术研究进展,系统分析了在不同环境下的栽培策略,包括实验室无宿主幼苗培育、盆栽成熟植株培育、大田规模种植以及野外定植等场景,整合了最新的盆栽实验结果,首次公开了田间预试验与野外互移栽实验的初步数据。研究揭示,要成功培育小米草,关键措施包括(1)利用低温层积法解除种子的休眠状态,(2)选择合适的宿主并控制其生长以防止竞争,(3)模拟自然生境中的良好排水土壤和全光照条件。尽管小米草属植物能在多种环境中生长,但在自然条件下培养出成熟的植株,仍需进一步开发更为可靠的标准化栽培方案。

1 引言
寄生植物是一个多样性丰富的类群,约含4,500个物种,其典型特征是具有称为吸器的寄生营养器官,可附着宿主植物窃取养分(Nickrent, 2020)。常见种类包括作物恶性杂草独脚金属植物(Striga, 列当科)、欧亚白果槲寄生(Viscum album, 檀香科)、草原野花鼻花属(Rhinanthus, 列当科),以及拥有自然界最大花朵的大花草(Rafflesia arnoldi R. Br., 大花草科)(Twyford, 2018)。寄生植物涵盖12个科(Westwood等, 2010),其栽培条件呈现高度异质性,即使近缘物种也可能需要截然不同的生长环境(Joel等, 2013)。这类植物的培育需克服若干特殊园艺难题:既要协调寄生物与宿主的双重需求,又需满足二者的互作条件。尤其多数寄生植物(特别是专性寄生种)需要宿主特异性信号诱导萌发(Yoneyama等, 2010),且几乎所有物种都依赖宿主才能旺盛生长(Albrecht等, 1999)。尽管如此,目前已有众多寄生植物被成功驯化(Pignone & Hammer, 2016)。近年随着生态修复领域对寄生植物的需求增长,其栽培技术与种子生产研究正引发广泛关注,相关技术规范的建立推广已成为当务之急。

小米草属目前记录约263个物种(Daniel Nickrent, 私人交流),广泛分布于南北半球温带地区及东南亚热带山区(Gussarova等, 2008),包含多年生与一年生类型(Yeo, 1973)。作为广谱性半寄生植物,虽具有光合自养能力且可独立生长,但与禾本科、非禾草或豆科等潜在宿主共生时表现更佳(Yeo, 1964; Brown等, 2020)。爱丁堡皇家植物园正以小米草为模式系统研究植物寄生机制,并探讨自然杂交、基因组进化及交配系统多样性等进化问题。不列颠群岛分布的20个本土种呈现出多维度表型变异,如生境偏好、伴生物种组成、二倍体与四倍体倍性共存(Yeo, 1956; Wang等, 2018)以及交配系统多样性(French等, 2005; Metherell & Rumsey, 2018)。野外调查证实该属存在广泛的种间杂交现象,可形成稳定的杂交后代种群乃至杂交起源的新物种(Stace等, 2015; Metherell & Rumsey, 2018)。

本文基于剑桥大学植物园分类学家Peter Yeo的开创性工作(其1956-1973年间发表了关于欧洲小米草分类与进化的系列论著),详细阐述英国本土小米草栽培技术的优化实践。Yeo的许多发现源自其栽培实验,包括种子育苗和野外草皮移植。本研究将系统讨论实验室、盆栽、大田及野生环境的全套实验栽培方案,结合不同宿主植物的共生栽培研究经验(Brown等, 2020, 2021; Becher等, 2020),并首次报道田间试验与野生互移植实验的初步结果。
2 小米草属植物栽培关键要点
当前植物学实验多选用易于栽培、株型矮小且生命周期短的物种作为研究对象,例如拟南芥(Arabidopsis thaliana)等模式植物(Wienkoop等,2010)。英国小米草虽同属一年生小型植物适于实验操作,但栽培条件更为特殊。其种子具有休眠特性,需经冷处理才能萌发(Yeo,1961;Liebst,2008)。这种季节性信号在自然环境中对确保种子春季同步萌发至关重要(Rubin & Friedman,2018)。通常采用两种破眠方法:冬季室外自然低温处理或冰箱强制冷诱导。后者可通过将种子置于无菌培养皿湿润滤纸上实现,仅需2-3滴水即可,过量易致霉变。受休眠限制,每年仅能培育一代,但在可控条件下或可缩短周期(见后文)。
小米草种子萌发率不稳定且普遍偏低(Yeo,1964)。野生种子的萌发成功率约40-50%,最低可至20%(Brown等,2020),其概率与种子质量直接相关。以北极小米草(Euphrasia arctica)小规模试验为例:20粒饱满种子中10粒萌发,中等质量种子6粒,皱缩种子则全军覆没(图1)。皱缩种子可能因发育缺陷、基因异常或母体资源限制而夭折(Stephenson,1981)。季末采集的种子萌发率通常最低,因多数健康种子已自然脱落。通过风选、筛选等清洁手段,或使用重力分选机处理大批量种子,可有效剔除劣质种子提高萌发率。贮藏前需将种子室温干燥一周,冰箱储存条件下(-4℃)可保持活力至少三年(Chapman等,2019)。户外盆栽实验中,多数种子集中在四月两周内同步萌发,但种间存在差异(图2)。
小米草萌发后需数日内寄生宿主方能健壮生长(Wettstein,1897;Yeo,1961)。其子叶肥大,下胚轴粗壮,但胚根相对短小(Simpson,1977)。根系扩展迅速,一周内即形成显著根毛。未能在两周内找到宿主者多死亡,少数植株地上部停滞生长但地下根系会持续扩张搜寻宿主(Yeo,1961)。宿主选择显著影响存活率:生长过快的宿主若无法被寄生,会与幼苗争夺光照导致后者死亡。总体而言,豆科或禾本科宿主的成活效果优于草本或木本植物。

图1 小米草种子质量的三种分类:(a)干瘪种子;(b)中等种子;(c)饱满种子。种子长度约2毫米。摄影:Hannes Becher
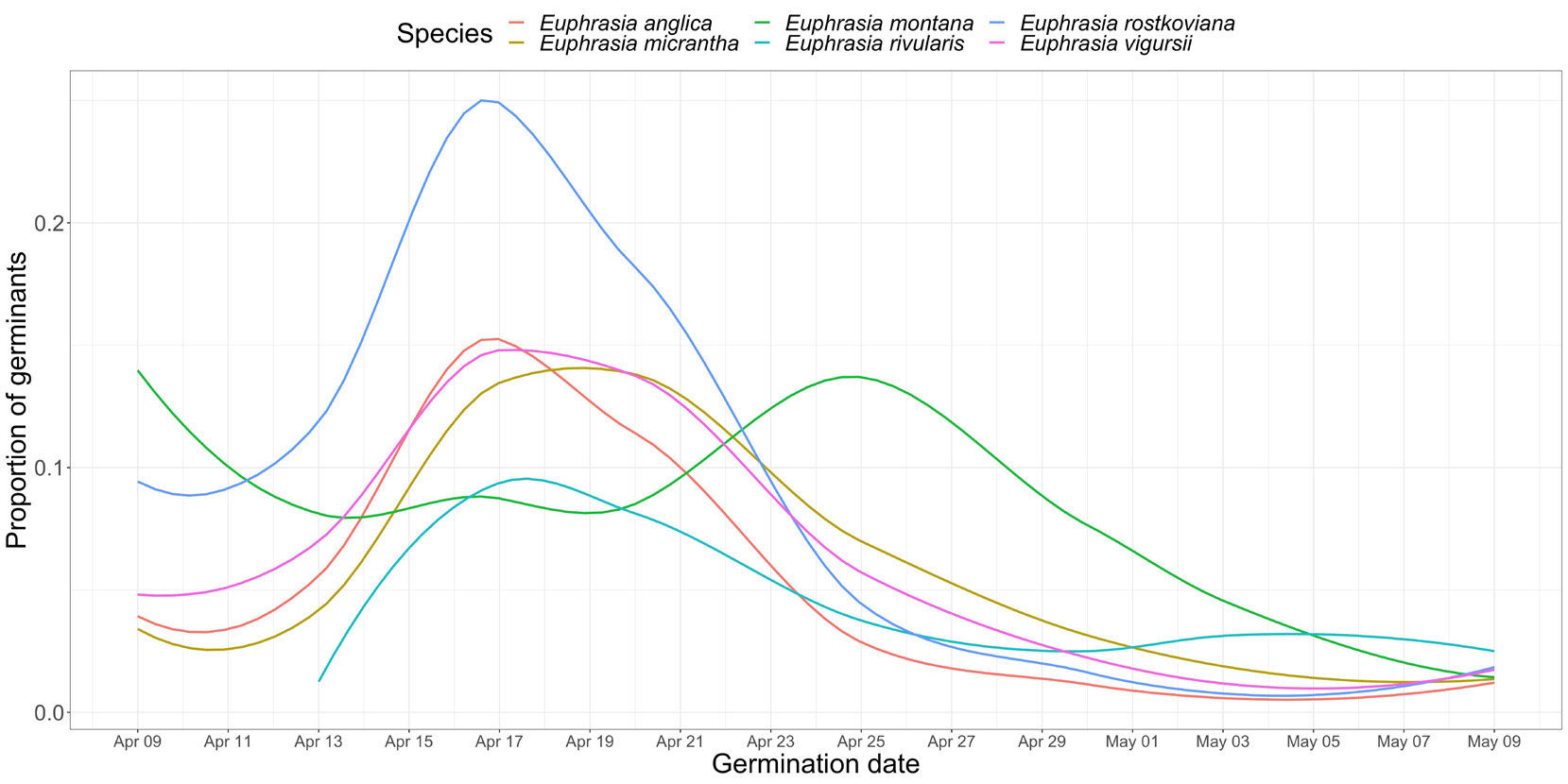
图2 爱丁堡皇家植物园小米草种子萌发情况。2018年春季对6种小米草进行每两日监测的发芽率数据(引自Brown等人2021年研究)
盆栽实验中,可在小米草种子萌发后小心引入寄主植物,以最大限度减少对幼苗的干扰。也可将寄主种子与小米草种子同期播种。但由于小米草发芽率不稳定且普遍较低(<50%),此方法会造成寄主植物大量浪费。此外,春季单独播种寄主植物可精准控制萌发时间,而秋季与小米草混播会导致寄主萌发与生长不同步,为对照实验引入干扰变量。更常规的做法是将小米草播种于现有植被中,但其竞争能力较弱,需确保植被低矮稀疏方能成功定植(见下文田间试验部分)。需注意的是,英国小米草均为一年生植物,所有个体在生长季结束时(通常持续至9月下旬)都会死亡。
无论在栽培环境还是野外,小米草都可能遭遇多种虫害。最严重的是蚜虫,主要侵害茎尖和叶片。严重感染时会导致叶片脱落,甚至造成植株死亡。喷洒皂液可缓解蚜虫危害,若为温室栽培,则需保持良好通风。另一常见病害是由鞘锈菌(Coleosporium,图3)引起的锈病,其转主寄主可能为松属植物(Ellis & Ellis, 1985),其病斑呈醒目的橙色,但对小米草的实际危害尚不明确。最后,波纹蛾属(Perizoma)某些鳞翅目幼虫会通过吐丝缀叶并啃食叶片侵害小米草(Fitter & Peat, 1994)。
3 实验室培养条件
为深入研究植物发育过程,常需在受控实验条件下培育植株。以小米草属植物为例,其种子可在无菌环境中的湿润滤纸上顺利萌发。操作时使用乙醇对培养皿和种子进行消毒,并用封口膜密封培养皿以防止污染。在萌芽前(无需补光),培养皿应置于4℃冰箱中保存。采用此方法成功培育野外采集的北极小米草种子,六周后即可观察到萌发现象。这种基于无菌培养皿的方法适用于获取幼苗,能满足特定实验需求(如细胞遗传学分析所需的幼根样本),但若用于培育成株则需进一步优化方案。
对于更精细的发育研究,可将植株培养于灭菌培养皿内的人工介质上。将种子置于1/4霍格兰氏营养液(一种广泛用于寄生植物培养的水培溶液)中,约10天后便可在4℃环境下萌发。

图3 小米草上的鞘锈菌(2020年9月摄于英国珀斯郡皮特洛赫里)。拍摄者:Hannes Becher
4 盆栽实验
采用盆栽法培育小米草的优势在于:相较于实验室人工培养基,天然基质能促成植物形成更自然的寄主互作关系。该方法特别适用于“同质园”研究(common garden studies),将不同来源不同物种的材料置于相同条件下培养。在统一条件下观察到的个体、种群或物种间表型差异,可归因于可遗传差异(即遗传或某些表观遗传因素,参见Riihimaki & Savolainen,2004)。若统一条件下的表型分化程度低于野生环境,则表明自然状态下的差异主要源于土壤、食草动物或可用寄主植物等环境因素。这类同质园实验被广泛应用于生态学、进化及遗传学研究领域,用以解析“先天与后天”效应的作用机制。
在爱丁堡皇家植物园苗圃完成了五项小米草盆栽同质园实验(表1)。所有实验均采用植物园树皮基质(有机材料混合而成,含50%细磨松树皮、35%木质纤维及15%堆肥绿植材料)进行露天播种育苗,前四项实验还在生长期将幼苗移入温室环境继续培养。
表1 爱丁堡皇家植物园苗圃小米草同质园实验概要
实验目的 | 小米草物种及其宿主 | 生长条件 | 参考文献 | |
1 | 探究不同物种形态学差异 | 11个物种的222个个体,均与三叶草宿主共同培养 | 2016年在玻璃温室中的盆栽实验研究 | Brown etal. (2020) |
2 | 研究宿主物种对小米草形态特征的影响 | 在8种不同的宿主物种上以及无宿主状态下,共记录了194个北极小米草个体 | 2016年玻璃温室中的盆栽实验 | Brown etal. (2020) |
3 | 量化不同宿主对小米草性能及存活率的影响 | 在45种不同宿主上1,379个北极小米草个体 | 2017年荷兰式玻璃温室盆栽实验研究 | Brown etal. (2021) |
4 | 探究小米草与宿主的相互作用 | 对13种宿主上6个不同小米草种群1,259个个体进行研究 | 2018年荷兰Venlo型温室的盆栽实验研究 | Brown etal. (2021) |
5 | 探究四倍体小米草属植物种间差异 | 不同来源的3种小米草物种的2个种群,每个种群分别与12种宿主物种共培养,共计2,124个个体 | 2019年室外框架内的盆栽实验 | Becher et al. (2020) |
初始实验旨在研究不同小米草属植物的生长模式,以及不同寄主和无寄主栽培条件对其的影响(Brown等,2020年)。研究表明:寄主种类会影响小米草的部分性状(如株高),但对其他性状(如开花节数)无显著影响;某些小米草物种(如亲缘关系较近的北极小米草、密序小米草Euphrasia confusa和林栖小米草Euphrasia nemorosa)在多数性状上存在重叠,而其他物种(如微花小米草Euphrasia micrantha与实验中的其他小米草)则表现出明显差异。第三项实验中,以北极小米草为研究对象,通过扩大寄主种类范围进行测试,发现不同寄主上的小米草存活率和适合度存在显著差异。2018年,首次系统测量了不同小米草物种在多种寄主上的适合度,以探究特定小米草-寄主组合间的寄生关系。第五项实验针对苏格兰设得兰群岛费尔岛隔离生境中的物种,研究其适应性与形态特征(Becher等,2020年)。通过标准化栽培草原型北极小米草、海岸型富拉小米草(Euphrasia foulaensis)及荒原型微花小米草,证实这些物种在同质花园中仍保持形态差异——尽管差异程度较自然环境下有所减弱。
所有五项实验均确保每个小米草种群-寄主组合至少设置30个盆栽,并预留约50%的发芽成功率。播种时将单粒种子置于盆土中央,这种布局有助于区分可能混杂的杂草种子。实验使用9厘米口径育苗盆,填装标准栽培基质。播种后覆以过筛细土,移置室外育苗架直至发芽(图4a)。寄主植物的精准接种对建立寄生连接至关重要:寄主种子于2月播撒于装有种植基质的育苗盘,4月将发芽不足两周的寄主幼苗移栽至含小米草的盆中。移栽植株需等距放置于小米草与盆器四角之一的中点位置。在群体共培实验中,该方法能保持寄主与小米草的标准间距,避免距离因素干扰。寄主接种过近可能导致过早附着(有利)或过度竞争(有害)。完成接种的盆栽被移入最终生长环境(温室或露天),夏季需定期修剪寄主以防遮荫及邻近盆栽根系蔓延,这对白三叶草等蔓生型植物尤为关键。
每月随机调整盆栽位置以降低区组效应(如某苗床植株长势偏优)。每日记录初花植株,所有性状测量均在初花当日完成(该标准化时点便于个体间比较),而适合度评估则贯穿整个生长季。

图4 爱丁堡皇家植物园苗圃用于小米草萌发的培育架,以及Becher等人(2020)在费尔岛开展的小米草实验。(a)(b)显示培育架上摆放20孔托盘,(c)(d)记录了2019年异常湿润的春季积水后土壤表面滋生的真菌。摄影:Hannes Becher

寄主选择对小米草健壮生长至关重要。若无寄主,小米草长势孱弱、植株矮小且难以开花(Brown等,2020)。试验初期常规使用三叶草作为寄主,但后续实验发现百脉根等豆科植物更佳(Brown等,2021)。长叶车前(Plantago lanceolata)是优良的阔叶寄主,冠毛犬尾草(Cynosorus cristatus)则属促进小米草生长的适宜禾本科植物。种源同样关键:商业种子遗传均一性高,能产生更整齐的生长表现;而野外采集种子虽可能导致生长参差,但更具生态代表性。费尔岛实验中同时采用了野生种子及野生帚石南(Calluna vulgaris)、欧洲刺柏(Juniperus communis)的扦插苗。使用多样性寄主(特别是喜酸性土壤的欧石楠类)时,可能需要调整栽培基质以优化寄主生长。
试验测试了多种培育环境:前两批实验使用老式Hartley木架温室,夏季高温导致植株死亡率较高且花芽脱落;随后两批改用新型连栋智能温室,其通风与气候控制系统更完善;第五批实验则全程在户外培育架(图4)中进行,金属围边有效防范动物与风害。户外培育虽能模拟野生状态下相对迟缓的生长节奏,但因框架遮阴导致长势较差,且2019年春季长期阴雨造成盆栽积水问题——这在可精准控水的温室内影响较小。未来计划采用遮阴极少的专用户外框架,并改良栽培基质排水性。
所有环境下均需适时补水。手动浇灌比自动灌溉更可靠,后者易造成水分不均。小米草偏好偏干环境,但这可能影响寄主生长。在贫瘠的小盆中,额外追肥对促进小米草生长必不可少:五月启动施肥计划,花期将频率从双周调至每周。使用注射器将液体肥料稀释至1.5%,通过细孔洒水器施用。追肥不仅改善寄主活力,还促进小米草生长并预防黄化。虽目前需肥量较大,但未来拟测试减量方案以更贴近自然土壤条件。
盆栽实验(尤其户外)需定期除草。季初小米草萌芽前重点清除杂草,避免其成为寄生对象。威胁植株的苔藓地衣也需剔除。所有除草操作均用镊子完成,以免伤及小米草或寄主根系。
5 田间试验
寄生植物通过抑制周边植被长势所产生的效应常被商业开发利用。小鼻花(Rhinanthus minor)作为常见寄生植物,广泛分布于多种自然与半自然植物群落中,并大量种植于草场以降低修剪维护需求(Westbury & Dunnett, 2007; Ameloot et al., 2005)。作为近缘半寄生植物,小米草属植物具有相似的潜在应用价值,该属物种广泛的生境适应性使其能在小鼻花不适配的区域发挥作用。
为实现规模化种植的种子生产,需开展小米草属的大田栽培。本研究与种子公司合作,通过设立田间试验探究其规模化栽培可行性。原计划采用成熟的小鼻花栽培方案,但早期尝试(通过划痕处理后向成熟草场播撒小米草种子的“小鼻花模式”)因禾草竞争而失败。故在2018/2019年度采用了改良方案:在两块24平方米的裸露地块上,以每平方米500粒(0.625克)的密度播种两个种源的北极小米草种子,并设置4种每平方米2克的不同寄主处理。所用寄主包括百脉根(Lotus corniculatus)、冠毛狗尾草(Cynosorus cristatus)、长叶车前(Plantago lanceolata)及草甸混合种(含17种以上草本与6种禾草)。试验期间仅实施最低限度管理,不进行灌溉,仅清除可能争夺资源的强竞争性杂草。
初步结果表明,大田栽培的北极小米草植株长势旺盛,株高至少达20厘米(图5)。此高度植株通常可产约300朵花,按每蒴果6粒可育种子计算,单株结籽量约1800粒。从收割效率考量,建议选用百脉根或长叶车前作为寄主——禾本科寄主会导致小米草茎基部弯曲缠绕,增加采收难度。其中长叶车前种子易与小米草种子分离,这种组合能有效保障种子纯度,这对种子生产至关重要。年际杂草防控将是栽培方案的关键环节,条播模式或具优势。通过该方案,北极小米草可实现大田规模化种植并获得理想种子产量。
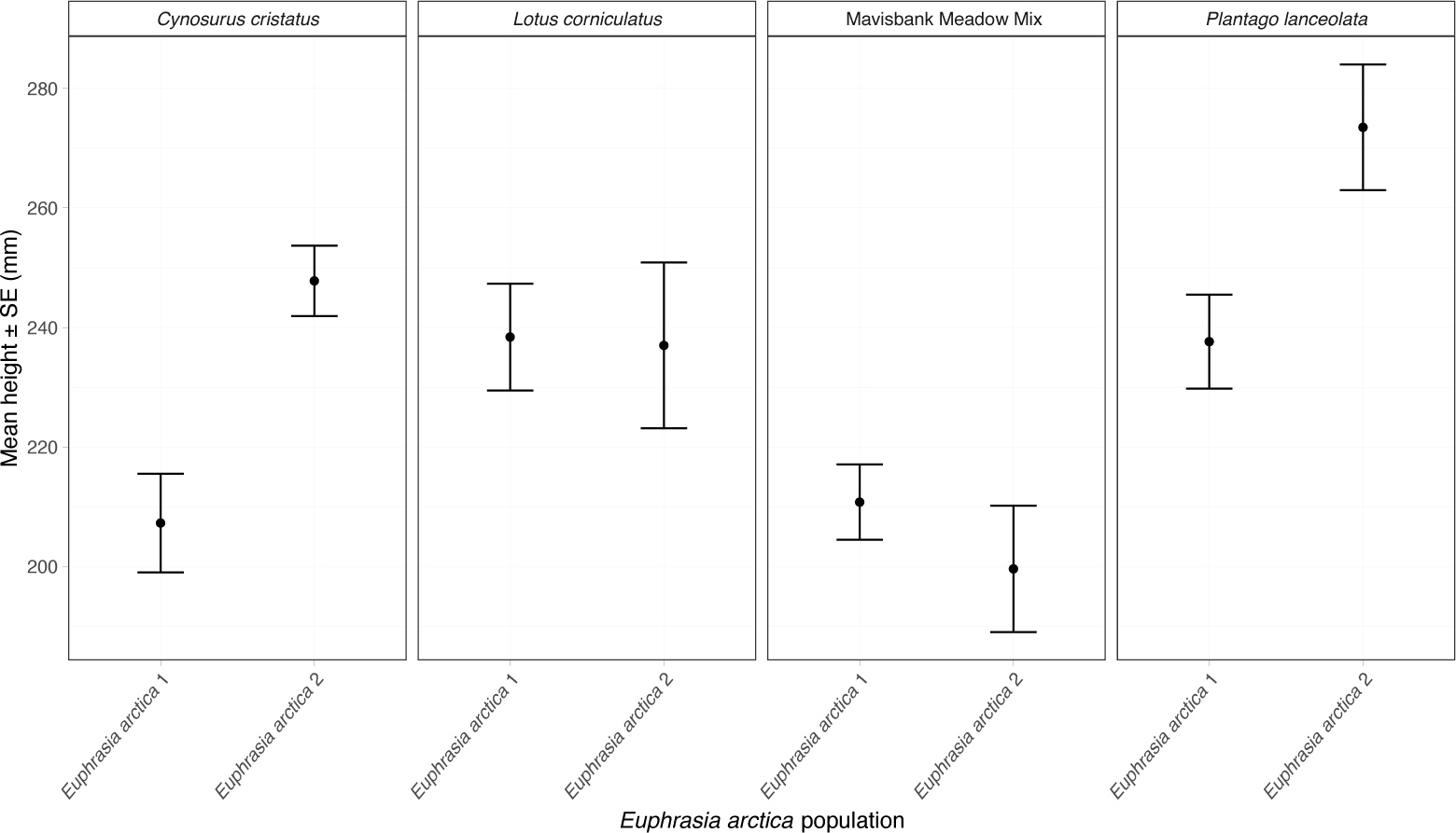
图5 在苏格兰安格斯试验田种植的北极小米草群体的平均株高及标准误。每子图显示植物在特定寄主上的生长状况。其中基因型1的数据整合自三个重复样方,基因型2(分别采集自苏格兰北贝里克和因弗基辛)的数据来自单一试验样方
6 互易移植实验
尽管前述方案模拟了自然条件,但在研究植物局部适应的进化生态学时,仍需进行野外栽培实验。局部适应是指植物在其原生地的表现优于其他地区(Blanquart等, 2013)。检验该现象的经典方法是互易移植实验:将来自不同地区的种群同时种植在彼此的原生地与异地进行对比观测(Kawecki & Ebert, 2004)。近期有学者采用此法研究了半寄生植物小鼻花(Rhinanthus minor)对海拔差异的适应性(Hargreaves & Eckert, 2019),证实该方法在半寄生植物研究中的可行性。针对费尔岛小米草开展了类似实验,通过与前述盆栽实验相结合,评估其对生境和寄主的局部适应性。
开展半寄生植物野外移植面临特殊挑战:一方面需要保留当地植被作为寄生宿主,无法彻底清除周边植物;另一方面必须防止土壤种子库中现存小米草的干扰。为此在种植穴中填充了经检验不含其他小米草种子的培养基质(确保根系可穿透以接触周围寄主)。在预实验(使用膨胀泥炭粒置于紧密契合的穴孔,见图6)未获成功——泥炭粒外包网层在生长季未降解,且多数发生位移(Becher个人观察)。最终在费尔岛的正式实验中,改用约翰英尼斯1号堆肥填充种植穴后播种。

图6:在因弗基辛(Inverkeithing)野外场地使用可扩展"Jiffy"育苗块培育小米草。(a)展示秋季排列的育苗块阵列,(b)显示春季幼苗的建植情况。埋入土中四个月后,育苗块外围网袋未见分解迹象。照片:Alex Twyford
本研究采用三种小米草各两个种群的种子,既用于爱丁堡皇家植物园的标准同园实验(参见Becher等,2020),也用于初步的交互移植实验。该移植实验旨在验证物种是否在其原生地的萌发表现优于新环境。在费尔岛(Fair Isle)设置了四个移植点:一处草地生境、一处石楠荒原生境及两处海岸生境。每个移植点设八个区块,每个区块包含72个种植穴,再细分为六个亚区(每亚区12穴),分别对应不同种源,小米草种子随机分配。次年五月的萌发成功率在不同生境和基因型间存在显著差异(表2)。
广义线性模型显示,原生地与移植地总体上对萌发率无显著影响(p=0.497),表明本实验中未发现萌发阶段存在局部适应的普遍证据。但各物种表现各异:草地的北极小米草在移植地的平均萌发率较原生地高10%(p=0.003),而海岸的富拉小米草则在原生地表现更优(差异12%,p<0.001)。微花小米草的萌发率差异不显著(p=0.468)。由此可见,小米草属植物的萌发响应存在种间差异,且呈现复杂模式。缺乏简单的局部适应信号(如原生地优势)并不完全出人意料,特别是考虑到实验空间尺度较小,以及生境差异的多维性(如土壤、暴露程度和植被等综合因素)。
虽然此类野外实验在多方面最能反映自然状态,但其执行面临诸多挑战。例如某些基质(如石楠荒地)难以挖掘、单株小米草的追踪非常困难,且自然条件下的萌发具有高度空间异质性。在费尔岛实验中,建植各阶段的自然损耗、周边植被竞争以及非实验来源小米草的本地萌发,使我们难以追踪植物的长期存活与生长。尽管如此,通过方法优化(试错法)、扩大样本量以确保植株回收率和统计效力,并实施精细监测,在自然环境开展此类实验仍具有可行性。
表2 费尔岛移植实验中小米草属植物的萌发情况
移植地点 | |||||
物种 | 原生种群地 | 威瑞维断崖Wirrvie Brecks | 布内斯 Buness | 观鸟点 Bird Observatory | 北灯塔 North Lighthouse |
北极小米草 | 菲尔教堂 FI Chapel | 34 | 30 | 17* | 35 |
学校 School | 24 | 18 | 21* | 32 | |
富拉小米草 | 南灯塔 South Lighthouse | 29 | 45* | 19 | 22* |
布内斯 Buness | 17 | 24* | 15 | 34* | |
微花小米草 | 威瑞维断崖Wirrvie Brecks | 24* | 25 | 16 | 29 |
简易跑道 Airstrip | 16* | 4 | 18 | 14 | |
表中数值表示每种基因型在每个位点的72个播种孔中的成功萌发数。星号(*)标注基因型在其原生栖息地种植的情况。使用R软件(R Core Team, 2013)建立二项分布广义线性模型进行统计分析,并采用emmeans包(Lenth, 2020)计算边际差异

7 结论与未来方向
小米草属是研究寄生进化、天然杂交重要性以及交配系统变异的理想模型。该属植物易于在实验室培养皿、温室或露天盆栽中栽培,且能寄生多种宿主植物,是探究寄生-宿主相互作用的绝佳体系。虽然商业化规模的田间试验已获成功,但野外互移植实验仍具挑战性,相关方案有待优化。小米草栽培的核心要点包括:
●种子层积处理对萌发至关重要,可通过人工模拟低温或露天越冬实现
●未经分选的野生种子萌发率约50%;夏末蒴果开裂前采集的种子经分选冷藏后萌发率最佳
●能显著促进小米草生长的优良宿主有限,主要为豆科植物及长叶车前、冠毛犬草等
●露天盆栽需全光照,温室栽培则需通风防暑以避免花芽败育
●在茂密草甸中,小米草竞争力弱于鼻花属,需清除现有植被以利建群
基于现有方案,未来研究可多向拓展:通过实验室栽培开展发育学研究,探究吸器形成及其与不同宿主的附着机制(参照鼻花属研究,见Rumer等,2007),并获得无污染组织样本用于基因组测序,揭示寄生相关基因(Yang等,2015);采用同质园实验探究地下寄生-宿主互作及宿主偏好性;开展大尺度互移植实验以解析本地适应性特征;建立生态修复中小米草定植方案及种子质量控制体系,为生态修复提供种源支撑。










延伸阅读:
Brown, M. (2021) “Student Project: Horticultural protocols for experimental studies of eyebrights (Euphrasia, Orobanchaceae)”, Sibbaldia: the International Journal of Botanic Garden Horticulture, (20), pp. 23–38. doi:10.24823/Sibbaldia.2021.319.
转载本文请联系原作者获取授权,同时请注明本文来自廖景平科学网博客。
链接地址:https://wap.sciencenet.cn/blog-38998-1481138.html?mobile=1
收藏