
 精选
精选

图1 被子植物(开花植物)
花的结构看似简单,其特点是具有确定的主轴,以严格的结构和时间顺序,在浓缩的同心区承载花器官,很少有物种逃脱这些限制。但如果放在适当的系统发生和发育背景下,那些背离这些限制的“不合群的”物种可以为花的进化史提供深入见解。在“我最喜欢的开花图片”中,邱园植物学家鲁道尔(Paula J Rudall)选择了两性霉草属(Lacandonia)花的纵剖面,这打破了花的雌雄蕊空间排列的基本规则:花是“由内向外”开放的,即心皮围绕着雄蕊,这种雄蕊在内雌蕊在外的模式在被子植物中几乎是独一无二的。当从其所属的科和目的背景下观察时,这个物种提供了比较进化领域许多迷人的植物科学兴趣。
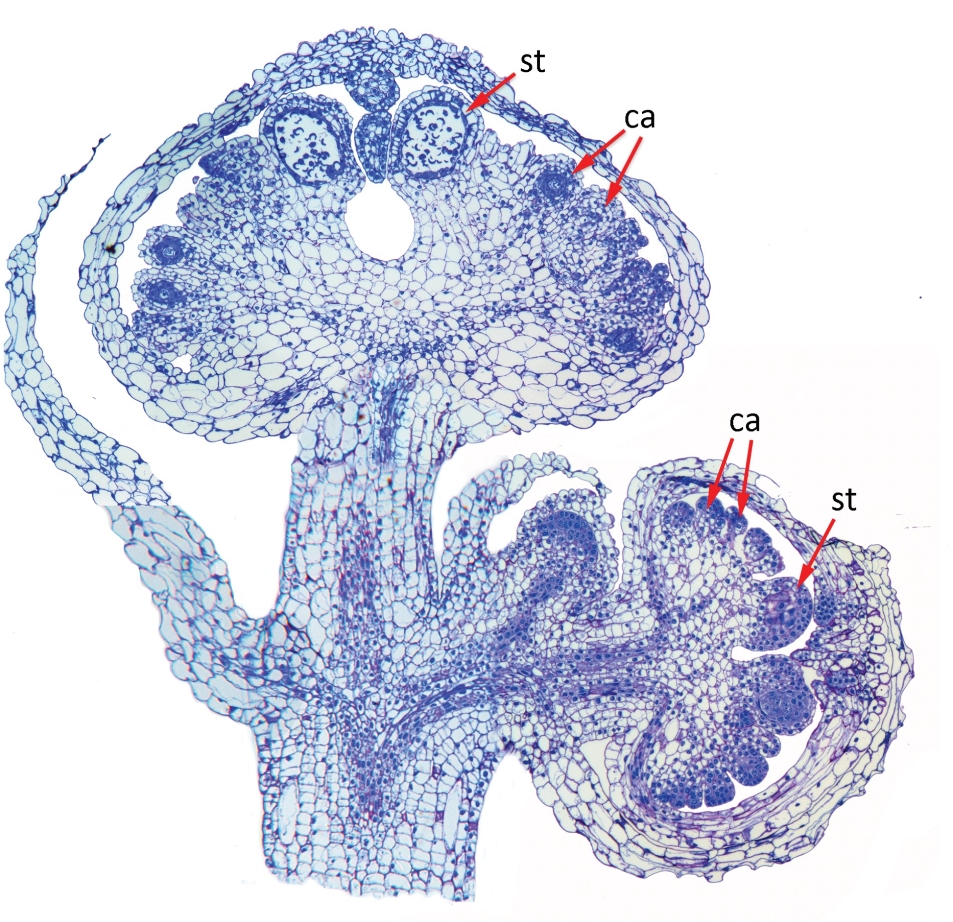
图2 巴西两性霉草花纵切片图:
巴西两性霉草(Lacandonia brasiliana)花序纵切面合成图,有处于三个不同发育阶段的花,显示出花的中心雌蕊雄蕊独特的排列方式,多枚雌性花器官心皮(ca)围绕在雄性花器官雄蕊(st)的周围,而不是通常所见的雄蕊围绕雌蕊周围。
被子植物的科,有时甚至是更高分类单元的整个目,往往可以通过特定的花特征来识别,即很容易辨认出一些主要植物类群的特征。这一事实表明,植物具有强烈的保持稳定发育程序特性。例如,包括现存被子植物物种 70% 以上的支系的一个相对一致的决定性特征是存在两轮五瓣花被,其中花的各部分交替轮生排列,每轮排列五个器官(Sauquet 等人,2017 年)。在非双子叶植物中,每轮五瓣的情况十分罕见,这凸显了在发育调控方面的重要性。与此相反,单子叶植物通常表现出三基数至五基数的状态,花的各部分三三两两地生在一起(Remizowa 等人,2010 年)。
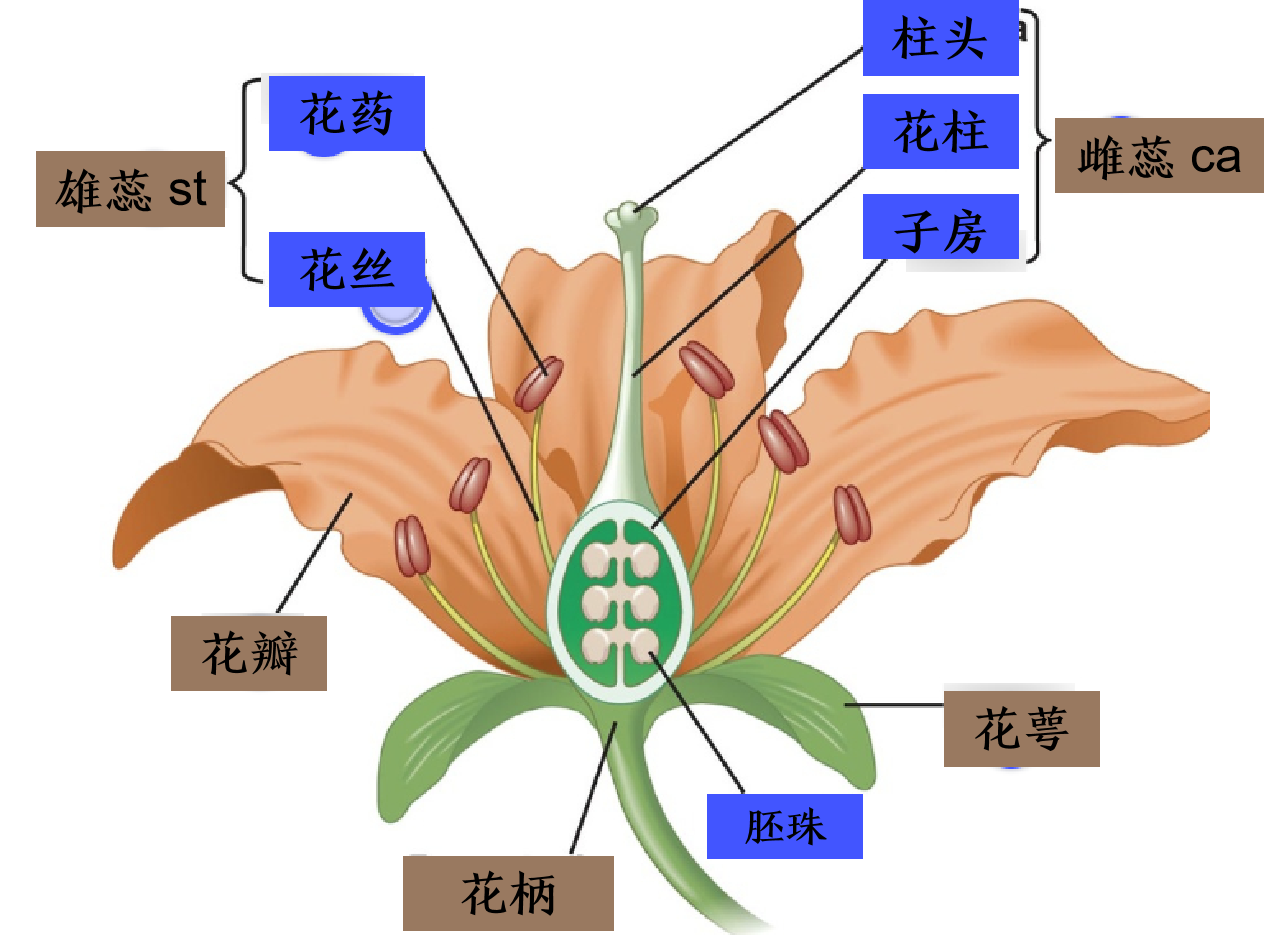
图3 被子植物完全花的构成图
在典型的被子植物花中,不育器官(花被)的外部区域围绕着花粉着生器官(雄蕊),而雄蕊又围绕着中央的雌性器官(心皮)。至少在拟南芥等模式植物中,这种器官排列的遗传基础在标志性的花发育 ABC 模型中得到了优雅的概括,激发了许多关于花结构和发育动态的新研究(Coen 和 Meyerowitz,1991 年;Bowman 和 Moyroud,2024 年)。令人惊讶的是,被子植物中几乎没有与这种模式相异的植物,这表明花器官的发育受到异常强烈的制约。但是,单子叶植物两性霉草属(Lacandonia)是一个明显的例外,它的花器官排列是相反的:数枚心皮围绕着早期形成的3枚中央雄蕊(Vergara-Silva 等人,2003;Ambrose 等人,2006)。两性霉草属的花器官倒置排花序在被子植物中几乎是独一无二的,这也导致一些学者质疑该物种的花-花序界限(Rudall,2003 年,2017 年)。B类和C类花同源基因的表达模式在两性霉草(L. schismatica)花中同样是倒置的(Álvarez-Buylla等人,2010年);这一观察结果凸显了ABC模型不可避免的局限性之一,该模型很好地解释了沿发育花轴基因的表达模式,但无法确定花轴本身的性质或进化起源。
霉草科(Triuridaceae)属于 “不合群”的单子叶植物露兜树目(Pandanales),该目中的其他几个类群都非常反常,缺乏可被轻易称为 “真正的”花结构;其中的例子包括环花草科环花草属(Cyclanthus)和露兜树科巨露兜树属(Sararanga)。这种不寻常的错位表明,在紧靠翡若翠科(Velloziaceae)之上的系统发育节点上,露兜树目在进化过程中失去了发育平衡(Rudall 和 Bateman,2006 年)。两性霉草属与其他霉草科植物的比较因缺乏完善的系统发育背景而受阻,这不仅反映了进化过程的复杂性,而且也是由于目前分子数据匮乏和该类群物种取样不足造成的。不过,现有的形态学比较数据表明,两性霉草属的花很可能是从一种具有更典型器官图案的结构演化而来的(例如 Nuraliev 等人,2020 年)。大多数其他霉草科植物的花都是单性的,不过也有少数物种的花是比较传统的两性花,如短柱三尾霉草(Triuris brevistylis),雄蕊与心皮的排列不是由内向外开放的(Rudall, 2003, 2008; Espinosa-Matías et al.)。
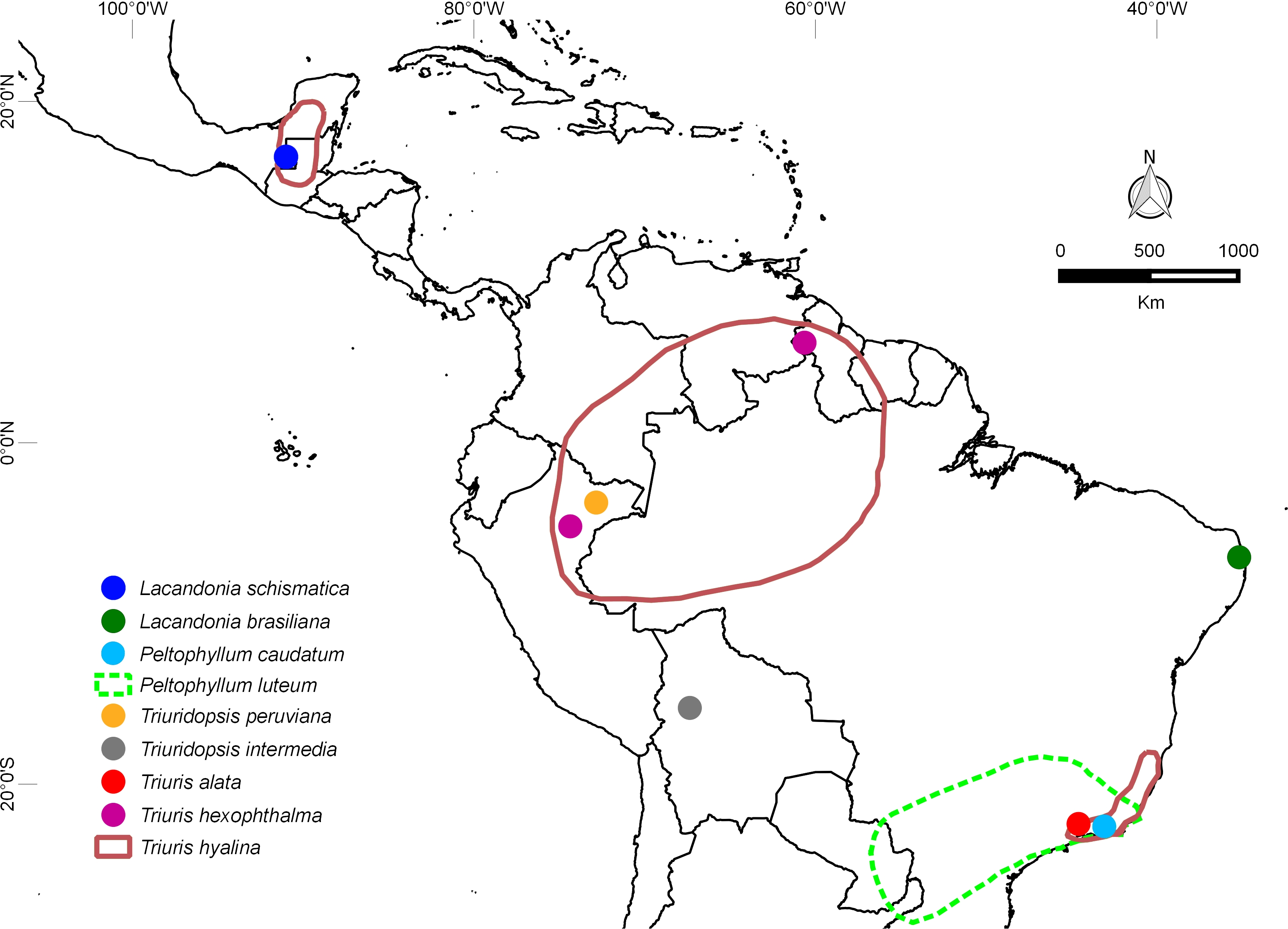
图4 两性霉草(蓝色点)和巴西两性霉草(绿色点)分布图
多年来,人们只知道两性霉草属的一个单一物种两性霉草(L. schismatica)(Martínez and Ramos, 1989),该种仅限于墨西哥东南部偏远的拉坎登雨林中稀有的零星种群。然而,最近巴西植物学家在第一个物种以南数千公里的巴西东北部发现了一种 两性霉草属新物种,他们将第二个物种描述为巴西两性霉草(Lacandonia brasiliana)(Melo 和 Alves,2012 年)。这一新发现促使科学家可以对这种花进行了更详细的研究,并将其作为露兜树目比较研究系列的一部分(Rudall 等人,2016 年)。两性霉草属的两个物种都出现在稀有而分散的种群中,这让科学家猜测这种奇特的形态是只进化了一次(可能是经过长途传播),还是在原地由两个独立的突变稳定下来形成了两个新物种。最近的研究表明,尽管 两性霉草属 的两个物种在地理上相距甚远,但它们在形态上确实非常相似。这表明,它们可能是由单个突变产生的,尽管这一推论很难得到确证。这两个物种的花都是由内向外开放的,中间有3枚雄蕊,周围有许多心皮;心皮本身排列在脊上呈放射状双排,被称为簇生,这是被子植物中另一个非常不寻常的特征。在这两个物种中,花药保持闭合状态,不会以正常方式散布花粉(Márquez-Guzmán 等人,1993 年;Rudall 等人,2016 年)。令人惊奇的是,在这两个物种中,受精和结籽显然是通过花粉粒在花药内萌发并通过同一植株的组织生长到雌性器官来实现的--这种罕见的现象仅在单子叶植物 浮叶慈姑(Sagittaria potamogetifolia)等少数其他开花植物中有所报道(Wang 等人,2002 年)。

图5 季节性半落叶林和巴西两性霉草
与霉草科的所有其他物种一样,两性霉草属也是一种菌根异养植物。由于叶绿素高度减少或缺失,因此它们无法通过光合作用产生碳水化合物;相反,它们依靠土壤中的真菌获得营养,因此是半透明的黄色而非绿色。因此,两性霉草属至少缺少两个支配植物结构的最重要因素:典型的花序和叶绿素介导的光合作用。这种与常规不同的显著组合会是巧合吗?很久以前,植物学家威廉‧赫姆斯利(William Hemsley,1843-1924 年)--曾任邱园标本馆馆长和查尔斯‧达尔文的好友--曾对霉草科的总体情况评论道:“这些小花在发育过程中似乎特别容易受到干扰”(Hemsley, 1907)。也许霉草科是绵状异养生物这一事实已经充分放宽了进化-发育的限制,从而允许 两性霉草属的花朵从内向外开放。最近的一项转录组分析表明,三个独立的髓营养型单子叶植物系的表达核基因发生了趋同的进化变化(Timilsena 等人,2023 年)。尽管这些结果仍未得出结论,但它们强调了与基因缺失相关的趋同进化的可能性。

图6 酒精保存的巴西两性霉草(A-E)和 六尾霉草(F)的花
然而,其他过程也可能导致物种克服强大的发育限制,进化出新颖的生长形式。例如,总状花序中的顶生花状结构可导致花状结构的次生衍生,如在早期分化的单子叶植物纲 泽泻目(Alismatales)中(Sokoloff 等人,2006 年)。由此产生的关键调控基因表达区的重叠可能会导致形态上的创新。唯一具有由内向外生殖单位的其他被子植物是睡莲的近缘植物 独蕊草属(Trithuria)(独蕊草科Hydatellaceae),它是一种不具有真菌异养短暂水生植物。与 霉草属一样,有关同源性的争论主要集中在 独蕊草科的花-花序界限上,该科在系统发育上靠近被子植物树的基部节点。独蕊草属 的生殖单位具有由内而外的浓缩结构,可能代表了由二次改造的花衍生出的类似花序的结构(Rudall 等人,2009 年)。
仅仅通过比较活体结构,即使结合相应的发育遗传学和系统发生学数据,似乎也不可能最终解决有关花的进化问题。形态学和分子的比较数据经常提供相互矛盾的证据。不过,这些研究表明,在花的进化方面仍有许多问题有待探索。对 霉草属 和其他此类 “不合群”植物的研究让我们更深入地了解植物结构的规则究竟是什么,以及这些规则是如何被定期打破的。

图7 浮叶慈姑 中国国家Ⅱ级濒危植物,北方特有的慈姑属物种
延伸阅读:
Rudall PJ (2024) My favourite flowering image: the remarkable inside-out flowers of Lacandonia. Journal of Experimental Botany, 75 (10): 2778–2780
转载本文请联系原作者获取授权,同时请注明本文来自廖景平科学网博客。
链接地址:https://wap.sciencenet.cn/blog-38998-1457115.html?mobile=1
收藏