博文
Nature | 线粒体: 嘿, 人! 该睡觉了欸_MedChemExpress
|
Section.01
人为什么会睡觉?
你有没有想过,我们为什么会睡觉? 我指的是......不受控的,打瞌睡。无论你凭借意志陪月亮熬到凌晨几点,终有一个瞬间闭眼投降。紧跟着的便是熬夜过后身体的种种不良反应,让你感觉身体被掏空,下一秒就要见到老天奶。。。
其实从进化的角度看,睡眠并不是件好事。因为如果生物体长时间无法感知和与环境互动,可能会增加被捕食的风险。但神奇的是!从秀丽隐杆线虫的简单神经系统,一直到人类大脑,睡眠从未被进化掉?![1]。
于是,科学家们将睡眠称之为一种非常特殊的现象。加之睡眠不足会带来认知能力受损、代谢功能障碍、患病风险增加,极端情况下甚至会导致死亡等有害影响。So,睡眠,既是动物界最常见的行为之一,也是自然界最大的谜团之一。
你可能要说了,存在即合理嘛~ 哎嘿!兄弟,它不仅合理,还有规律——生物钟。
生物钟,又称昼夜节律 (Circadian rhythm):是一种生理现象,以内源性、持续的,呈现以约 24 小时为周期的变动。
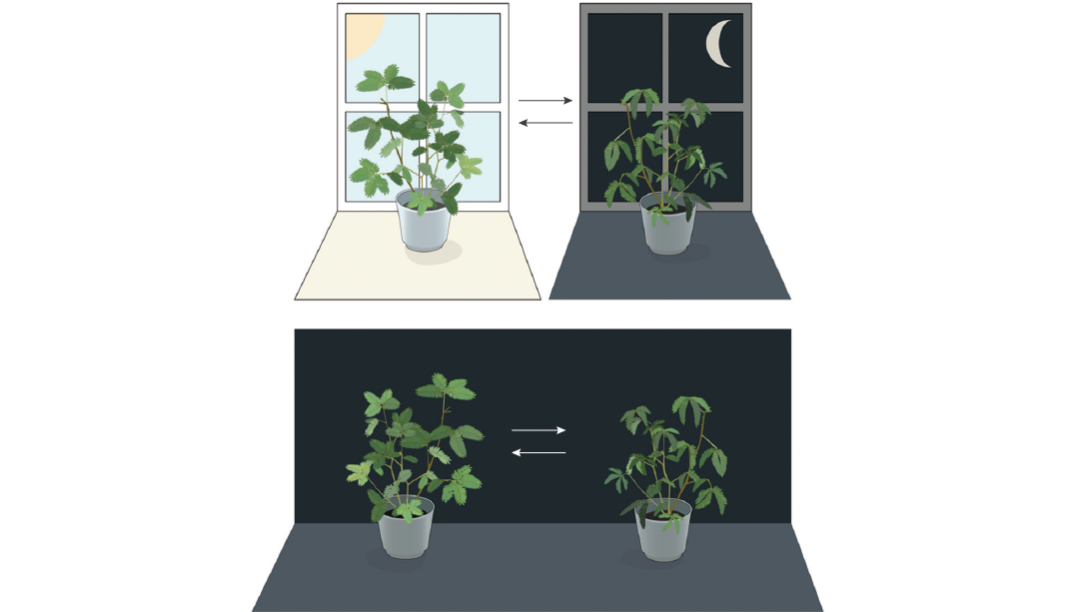
图 1. 含羞草的叶片白天张开,黄昏时闭合。且当含羞草处于持续黑暗中时,其叶片仍会维持正常的昼夜节律[2]。
早在 18 世纪,天文学家就发现植物具有内源性的生物节律。为了了解这种内源性昼夜节律的机制,科学家做了诸多研究。直到多年后,3 位美国科学家杰弗理•霍尔 (Jeffrey C. Hall)、迈克尔•罗斯巴希 (Michael Rosbash)、迈克尔•杨 (Michael W. Young) 揭示了果蝇生物钟的调控机制,也因此荣获 2017 年的诺贝尔生理学或医学奖。科学家成功克隆了 "周期" 基因 period,period 基因的编码蛋白 PER 夜间积累,白天降解,其蛋白水平的 24 h 周期性波动与昼夜节律同步。TIM 蛋白可以与 PER 蛋白结合,携带后者进入细胞核,通过抑制 period 基因活性来完成负反馈调控,减少自身蛋白的合成。此外,基因 doubletime 编码的蛋白 DBT 可以延缓 PER 蛋白的积累,使后者的震荡频率基本吻合 24 h 周期 (图 2)。
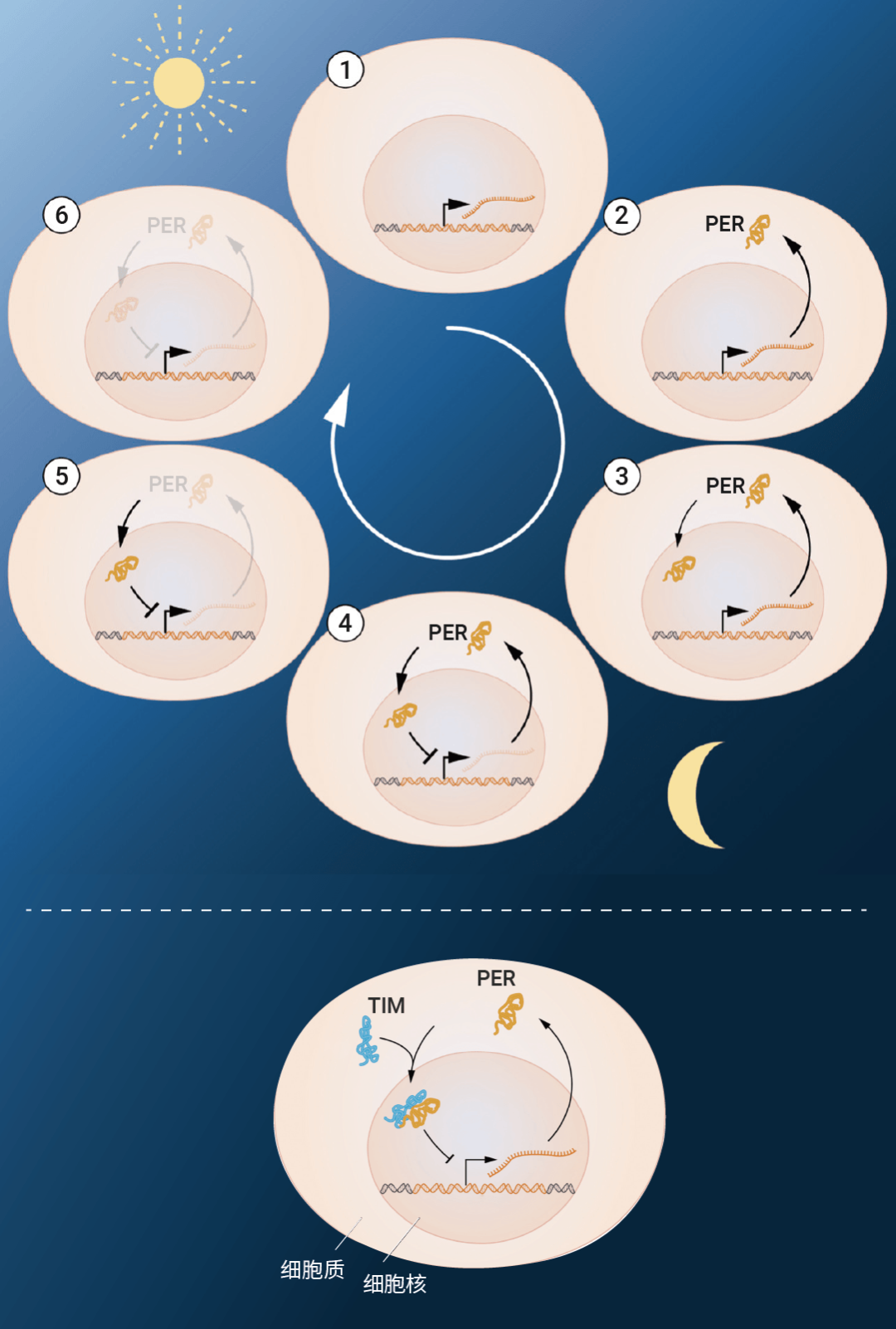
图 2. period 基因负反馈调控环路[2]。
2017 年的诺贝尔生理学或医学奖也让果蝇这一卓尔不凡的小动物再次身显名扬,这也是第五座因果蝇而获得的诺奖。
果蝇,作为研究睡眠相当有用的动物模型[3]。研究发现,在其睡眠调节神经回路中,背侧扇形体 (dorsal fan-shaped body, dFB/dFNBs) 是主要的促进睡眠区域[4]。近期 Nature 发表的研究性论文发现——当在背侧扇状体 (dFBNs) 中检测到过多的线粒体损伤时,就会触发睡眠。
Section.02
Nature 发文:
线粒体起源的睡眠压力
2025 年 7 月 16 日,Nature 发表标题为“Mitochondrial origins of the pressure to sleep”的研究性论文,该论文研究了睡眠压力 (sleep pressure) 的线粒体起源,探讨了大脑中分子变化与睡眠需求的关联[5]。
线粒体基因表达上调
首先,研究人员通过分析休息和睡眠剥夺的果蝇的单细胞转录组,发现在睡眠剥夺后,上调的转录本主要在投射到背侧扇状体 (dFBNs) 的睡眠控制神经元中表达,且在 dFBN 中上调的转录本,几乎只编码参与线粒体呼吸和 ATP 合成的蛋白质 (图 3a)。
如图所示,通过对 12 小时夜间睡眠剥夺后 dFBN 中水平发生变化的 122 个转录本进行基因本体分析,发现睡眠剥夺导致编码电子传递复合物 I-IV、ATP 合酶 (复合物 V)、ATP-ADP 载体 sesB 和三羧酸循环酶 (柠檬酸合酶 kdn、琥珀酸脱氢酶 B 亚基和苹果酸脱氢酶 Men-b) 成分的转录本选择性上调,而参与突触组装、突触囊泡释放和突触前稳态可塑性的基因产物则选择性下调。
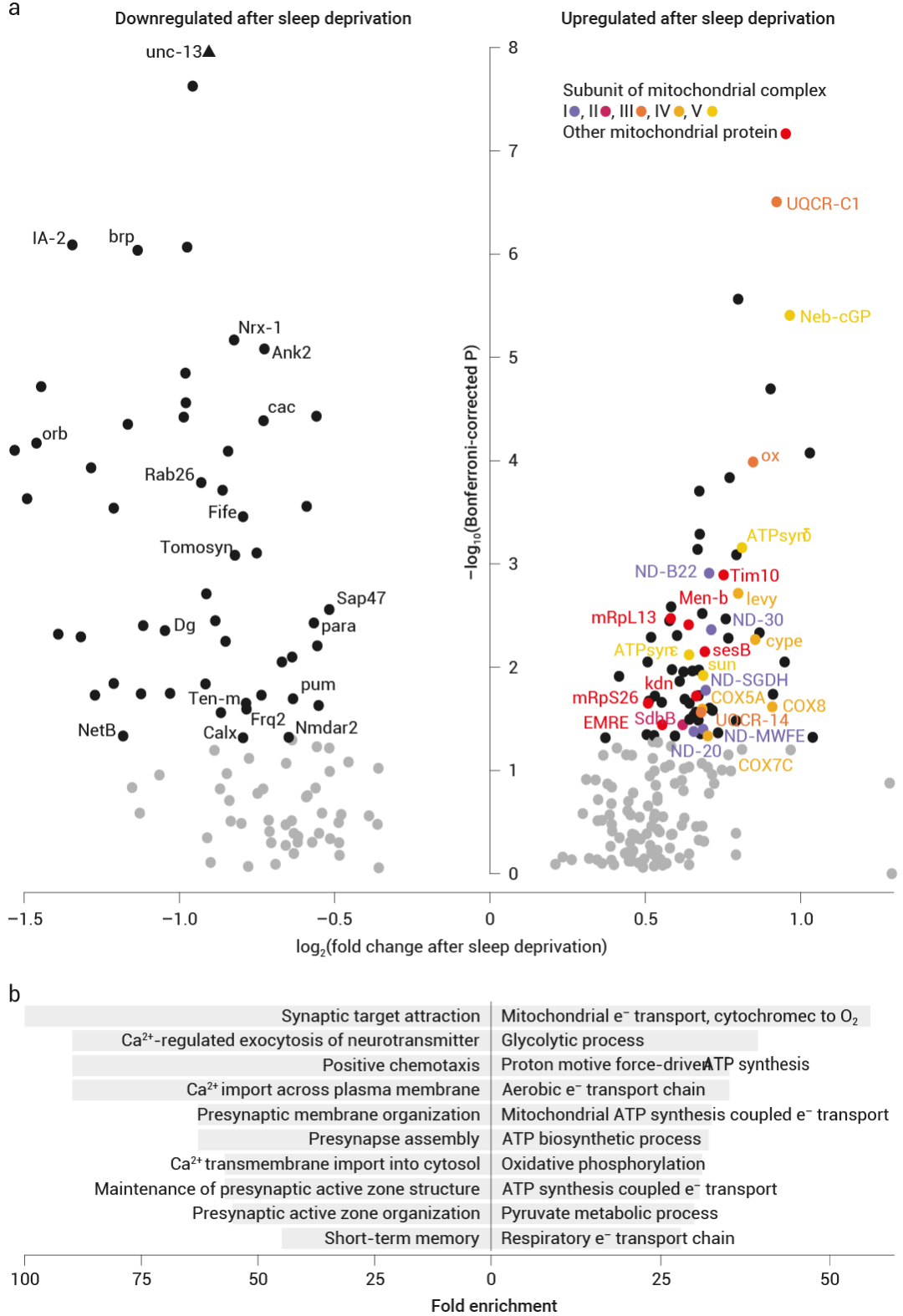
图 3. 线粒体成分在 dFBN 对睡眠剥夺的转录反应中的突出作用[5]。
睡眠剥夺 — 线粒体: 不睡么? 我要 "裂" 了
作者用基质定位的绿色荧光蛋白 (mito-GFP) 标记了 dFBN 的线粒体,并在一系列独立的实验中,用共聚焦激光扫描显微镜 (CLSM) 或光学光子重排显微镜 (OPRM) 对神经元的树突区进行成像。发现睡眠剥夺后的基因表达变化伴随着线粒体碎片化、线粒体自噬增强以及线粒体与内质网之间接触数量的增加,恢复睡眠后,形态变化是可逆的。
首先,在线粒体形态上,研究发现无论是机械搅拌还是人为提高多巴胺水平所致的睡眠剥夺,都会导致 dFBN s 中线粒体的尺寸缩小、伸长率降低、分支减少 (图 4a-b)。并导致动力蛋白相关蛋白 1 (Drp1) (外膜的关键裂变动力蛋白) 从细胞质移位到线粒体表面 (图 4c),OPRM 检测到线粒体数量的伴随增加 (图 4b),表明线粒体发生分裂。同时,睡眠剥夺的果蝇的 dFBN 中接触位点计数较高,也就是说睡眠剥夺导致 dFBN 中线粒体-内质网接触更加丰富,从而形成补充过氧化脂质的通道 (图 4d)。同时,mito-QC (一种检测线粒体进入酸性自噬溶酶体的比例传感器) 则报告了线粒体自噬增强 (图 4e)。而在随后的恢复睡眠期间,线粒体的增殖和融合导致它们的体积、形状和分支长度反弹至基线值以上 (图 4a-b)。这表明线粒体损伤可能会引发睡眠冲动。
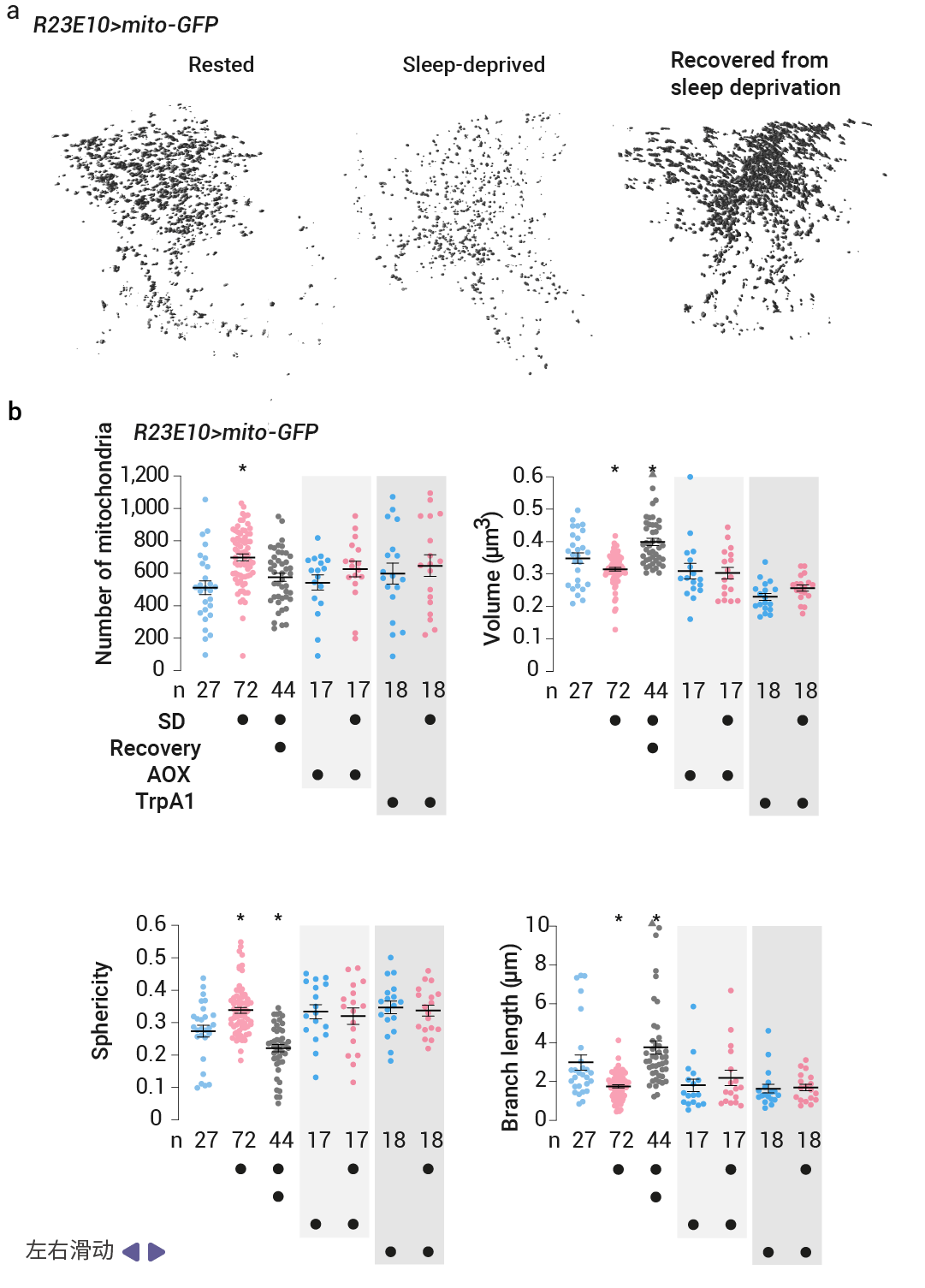
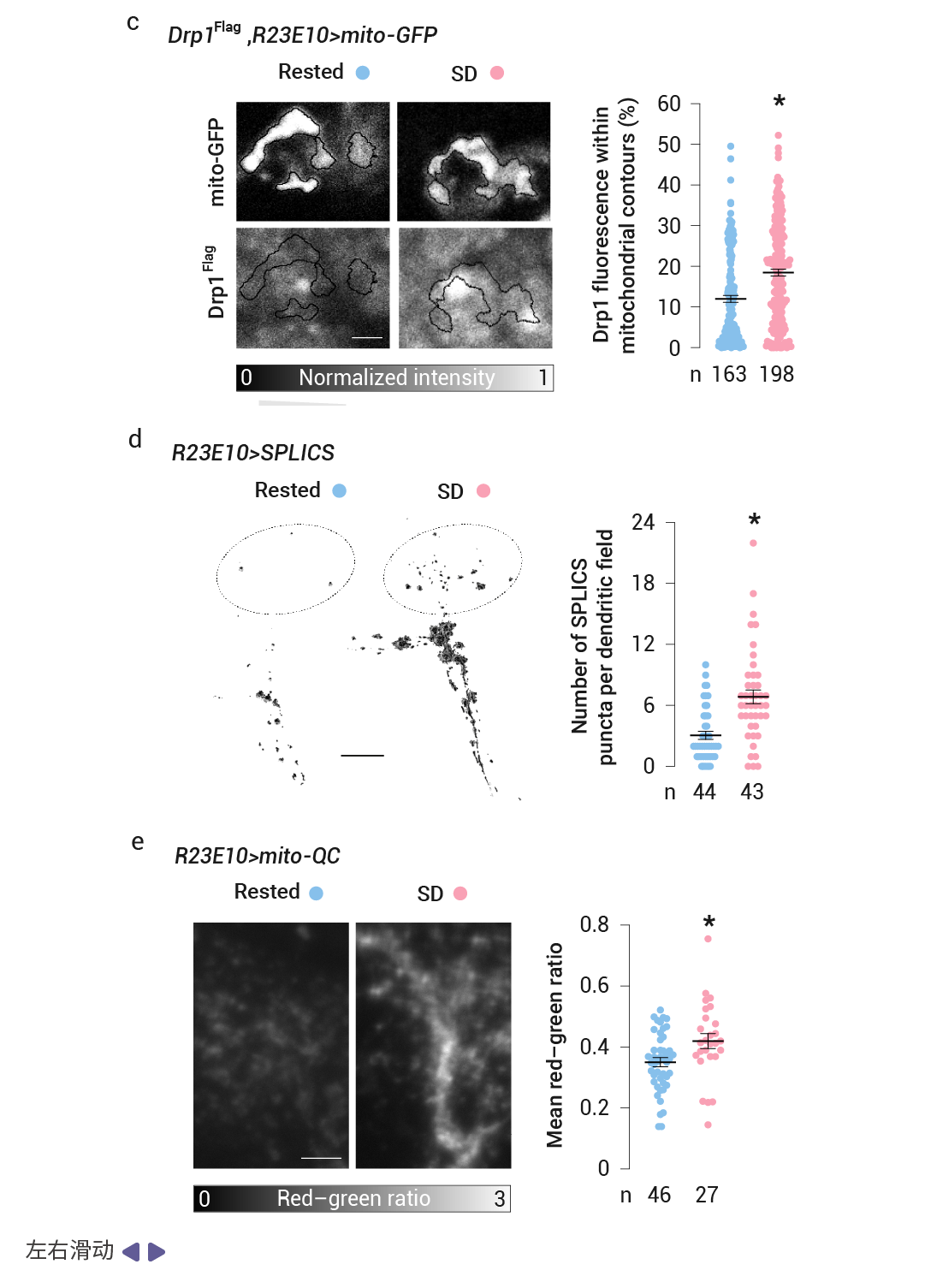
图 4. 睡眠改变线粒体的动态变化[5]。
a-b. 休息的苍蝇、睡眠不足的苍蝇和睡眠不足 24 小时后恢复的苍蝇的 dFBN 树突的 OPRM 图像堆栈中自动检测到的线粒体的体积渲染 (a) 和形态参数 (b)。c. Drp1 募集。通过表达 R23E10-GAL4 驱动的线粒体-GFP(顶部)和来自内源性基因座的 Drp1 Flag (底部)的果蝇的dFBN 体细胞的单共聚焦图像平面。睡眠剥夺会增加自动检测到的线粒体轮廓内的细胞抗 Flag 荧光的百分比。d. 线粒体-内质网接触。e. 线粒体自噬。
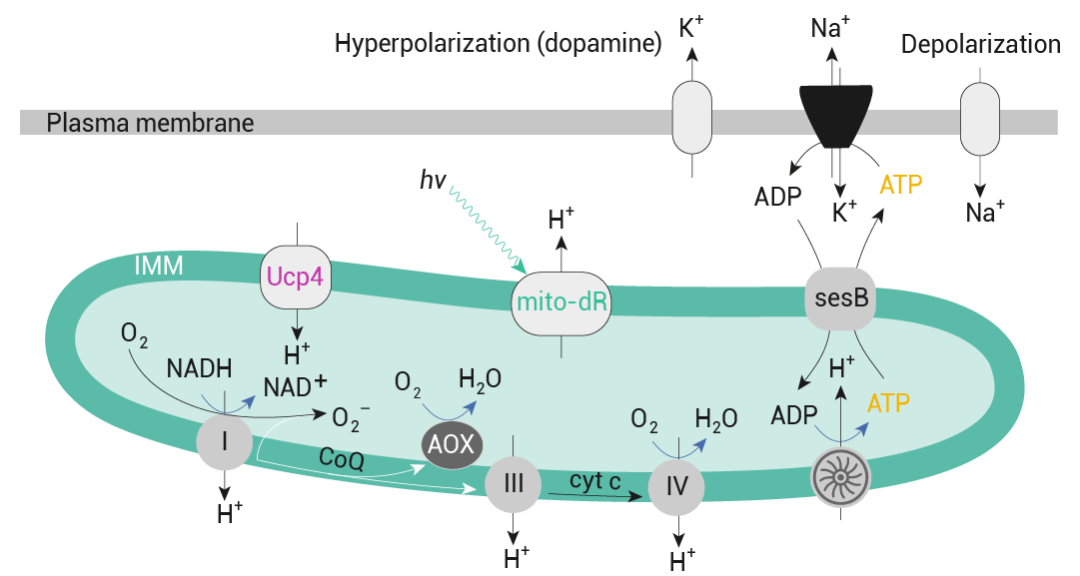
图 5. dFBN 通过将诱导睡眠的脉冲放电与线粒体呼吸相结合来调节睡眠[5]。
线粒体:裂变—睡的少,融合—叫不醒
随后,作者人为地人为控制了线粒体的裂变和融合。因为如果线粒体裂变和融合之间平衡的变化是反馈机制的一部分。那么在 dFBN 中实验性地诱导这些稳态反应应该会改变睡眠的设定点:线粒体的分裂预计会减少睡眠时间和深度,而反之线粒体融合预计会增加睡眠时长和深度。
于是,作者对在线粒体动力学中具有中心调控作用的三种 GTP 酶进行了实验控制 (图 4a):(1) 裂变动力蛋白 Drp1,以及内外线粒体膜的整合蛋白 (2) 视神经萎缩 1 (Opa1) 和 (3) 线粒体融合蛋白子 (Marf) (它们在顺式和反式的聚合使得相应的膜融合)。
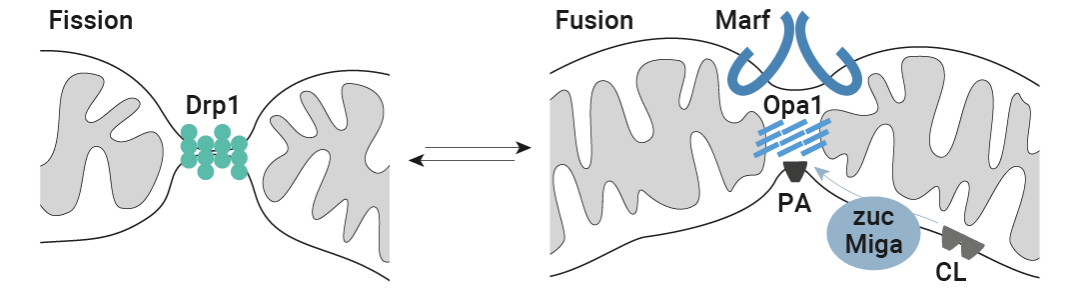
图 6. 线粒体裂变 (绿色) 和融合机制 (蓝色)[5]。
通过过表达 Drp1 或 RNAi 介导的 Opa1 减少 (在较小程度上也包括 Marf) 来使 dFBN 线粒体碎片化,减少了睡眠 (图 7a-b)。消除对睡眠剥夺的稳态反应 (图 7c),并且无论睡眠史如何,均降低了 dFBN 中的 ATP 浓度。
将平衡倾向于线粒体融合则产生了相反的效果:在 dFBN 中敲低 Drp1 或过表达 Opa1 加 Marf——或者单独过表达 Opa1,但不包括单独过表达 Marf——增加了基线及反弹睡眠 (图 7a-c),并提高了觉醒阈值,且不会引起过表达伪影或明显的发育缺陷。
报道显示,在泛神经元或胶质细胞 RNAi 敲低 Drp1 或 Marf 基因后,睡眠丧失现象普遍存在。在 dFBN 中,Drp1 或 Marf 基因的表达会双向改变睡眠 (图 7a-c)。
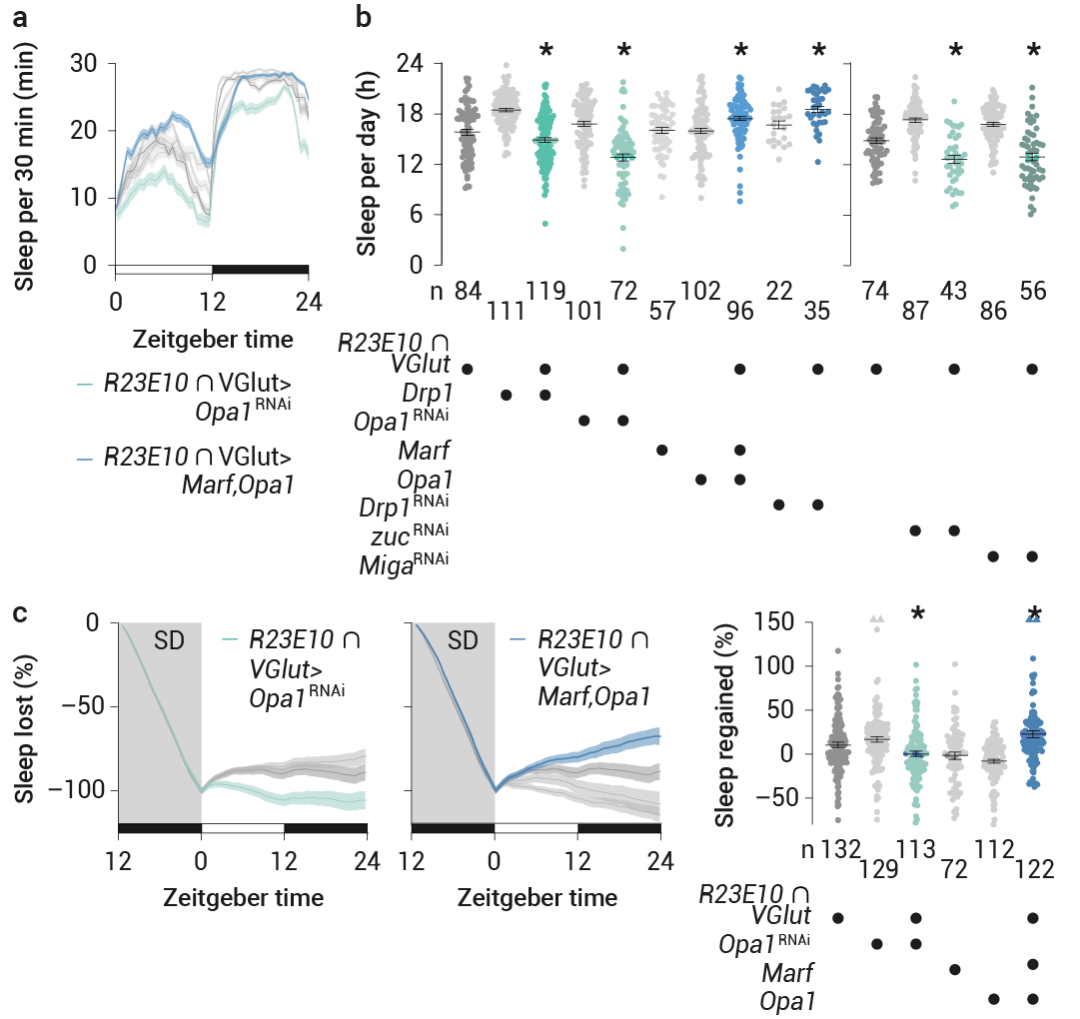
图 7. 线粒体动力学改变睡眠[5]。
最后,作者提到睡眠可能是为了满足代谢需求而进化的。比如,线粒体融合后,AgRP 神经元的电输出会增加,从而促进体重增加和脂肪沉积;这就像线粒体融合后 dFBN 的电输出会增加以促进睡眠一样。删除 AgRP 神经元中的线粒体融合蛋白会损害食物摄入,就像干扰 dFBN 中的线粒体融合会损害睡眠诱导一样。这些相似之处表明,睡眠压力和饥饿感都源自线粒体,并且电子会流经各自反馈控制器的呼吸链,决定何时需要恢复平衡。
产品推荐
是选择性的发动蛋白相关蛋白 1 (Drp1) 抑制剂。Mdivi-1 是一种有效的线粒体分裂/线粒体自噬 (mitophagy) 抑制剂。
是一种线粒体融合 (mitochondrial fusion) 的小分子抑制剂。
MitoTracker Deep Red/ MitoTracker Green (HY-D1783)
线粒体红色/绿色荧光探针。
是一种广泛用于检测线粒体膜电位的理想荧光探针。正常线粒体内,JC-1 聚集在线粒体基质中形成聚合物,聚合物发出强烈的红色荧光 (Ex=585 nm, Em=590 nm) ;在线粒体膜电位较低时,JC-1不能聚集在线粒体的基质中,产生绿色荧光 (Ex=510 nm, Em=527 nm) 。
是一种针对线粒体的荧光染料,能特异性靶向线粒体。TPE-PY 主要用于生物成像领域,特别是活细胞中线粒体的特异性染色研究,有助于观察线粒体的位置、形态和数量。

[1] Ly S, Pack AI, Naidoo N. The neurobiological basis of sleep: Insights from Drosophila. Neurosci Biobehav Rev. 2018 Apr;87:67-86.
[2] 2017年度诺贝尔生理学或医学奖, 生命科学, 2017 年 12 月, 第 29 卷 第 12 期, DOI: 10.13376/j.cbls/2017161
[3] Tainton-Heap LAL, et al. A Paradoxical Kind of Sleep in Drosophila melanogaster. Curr Biol. 2021 Feb 8;31(3):578-590.e6.
[4] Kato YS,et al. Interneurons of fan-shaped body promote arousal in Drosophila. PLoS One. 2022 Nov 21;17(11):e0277918.
[5] Kato YS, et al. Interneurons of fan-shaped body promote arousal in Drosophila. PLoS One. 2022 Nov 21;17(11):e0277918.
https://wap.sciencenet.cn/blog-3536222-1496419.html
上一篇:ATPγS tetralithium salt,93839-89-5,≥98.0%_MedChemExpress
下一篇:干货分享 | DSS 诱导结肠炎: 不同分子量差异有哪些?选对才能造好模!_MedChemExpress