ВЉЮФ
[зЊди]ЁОNature Communication IF=17ЁПжмЦкадзищЕѕЃЛЏЕїНкаЁЪѓЕФ TLR9 аХКХДЋЕМКЭздЩэУтвп
|||

Toll бљЪмЬх ЃЈTLRЃЉ дкЦєЖЏЯШЬьУтвпЗДгІжаЦ№зХживЊзїгУЁЃЫфШЛДѓЖрЪ§ TLR ЮЛгкжЪФЄЩЯЃЌЕЋ TLR3ЁЂTLR7ЁЂTLR8 КЭ TLR9 ПЩЪЖБ№ФкЬхжаЕФжТВЁКЫЫсЁЃетаЉФкЬх TLR ашвЊАщТТЕААзUNC93B1ВХФмЮШЖЈКЭДгФкжЪЭј ЃЈERЃЉ-ИпЖћЛљЬхЕНФкШмУИЬхЕФдЫЪф.вЛЕЉНјШыФкЬхЃЌетаЉ TLR ЕФАћЭтНсЙЙгђБиаыБЛФкыФУИЧаИюЃЌШЛКѓ TLR ВХФмгааЇЕиИажЊКЫЫсЁЃФкЬхTLRЕФВЛЕБдЫЪфЕМжТаЁЪѓбЯжиЕФздЩэУтвп.ç¨дЬхбмЩњЕФКЫЫсЭтЃЌTLR7ЁЂTLR8 КЭ TLR9 ЛЙПЩвдЪЖБ№ФкдДадКЫЫсЃЌВЂгыздЩэУтвпадМВВЁгаЙиЃЌР§ШчвјаМВЁЁЂЯЕЭГадгВЛЏжЂКЭЯЕЭГадКьАпРЧДЏ ЃЈSLEЃЉЁЃ
дкSLEжаЃЌTLR7ЕФRNAИагІКЭTLR9ЕФDNAИагІЪЧМВВЁЗЂВЁЛњжЦЕФЙиМќЃЌЕЋЧБдкЕФЗжзгЛњжЦВювьКмДѓЁЃЛюЛЏЕФНЌЯИАћбљЪїЭЛзДЯИАћ ЃЈpDCЃЉ БэДяИпЫЎЦНЕФ TLR7 КЭ TLR9ЃЌВЂЗжУкДѓСП I аЭКЭ III аЭИЩШХЫи ЃЈIFNЃЉЁЃетжжПьЫйЯИАћвђзгЗДгІЪЧгЩ pDC жа IRF7 КЭ NF-ІЪB ЕФзщГЩаЭБэДяНщЕМЕФЃЌЖј NF-ІЪB ЗжзгЭООЖЪЧЦфЫћУтвпЯИАћЃЈШчОоЪЩЯИАћЃЉжаЕФжївЊЗДгІНщжЪЁЃpDC ДњБэСЫЪїЭЛзДЯИАћЕФЖРЬиЦзЯЕЃЌашвЊзЊТМвђзг E2-2 РДЗЂг§КЭЮЌГжЦфЩэЗнЁЃpDC ПЩвдЭЈЙ§ФкЬх TLR7 КЭ TLR9 БЛздЩэКЫЫсМЄЛюВЂВњЩњДѓСП IFN-I31.ЗДЙ§РДЃЌpDCбмЩњЕФIFN-IПЩвдМЄЛюЫшЯЕЯИАћЃЌжБНгЕїНкTЯИАћКЭBЯИАћЕФЗжЛЏЁЃШЫРрКЭаЁЪѓФЃаЭжаЯЕЭГадКьАпРЧДЏЕФЗЂВЁЛњжЦвдЙ§ЖШЕФIFN-IаХКХДЋЕМЮЊЬиеїЁЃpDC жа IFN ВњЩњЕФЪЇЕїгажњгкЖржжздЩэУтвпадМВВЁЃЈШч I аЭЬЧФђВЁЁЂвјаМВЁЁЂЯЕЭГадгВЛЏжЂКЭ SLEЃЉЕФжТВЁРЉді14,36ЈC39.дкаЁЪѓжаЃЌpDCШБЗІПЩгааЇдЄЗРЯЕЭГадгВЛЏжЂКЭSLEЕФЗЂВЁЛњжЦЁЃвђДЫЃЌpDCsвьГЃВњЩњIFN-IЪЧздЩэУтвпЕФживЊЧ§ЖЏвђЫиЁЃ
S-зищЕѕЃЛЏЪЧвЛжжПЩФцЕФЗвыКѓаоЪЮЃЌЦфжаАыызАБЫсВаЛљЙВМлСЌНгЕНБЅКЭ C16 жЌжЪзищЕЫсѕЅЩЯЁЃзищЕѕЃЛЏзДЬЌгажњгкЕААзжЪНсЙЙКЭЮШЖЈадЁЂФвХндЫЪфКЭФЄУЊЖЈЁЃDHHCЕААзѕЃЛљзЊвЦУИЕФвЛДѓМвзхПЩвдS-зищЕЫсЛЏИпЖћЛљЬхЁЂАћжЪШмНКЛђЯИАћФЄжаЕФЕААзжЪЁЃМИжжРраЭЕФЫЎНтУИЃЌШчѕЃЛљЕААзСђѕЅУИКЭзищЕѕЃЕААзСђѕЅУИЃЌПЩвдЗжБ№ЪЙАћжЪШмНКЛђШмУИЬхжаЕФЕААзжЪШЅзищЕѕЃЛЏЁЃдкУтвпЯЕЭГжаЃЌзищЕѕЃЛЏбЛЗвбБЛжЄУїЖдМИИіЙиМќЕФЯИАћЭЈТЗКЭУтвпЙ§ГЬЃЈШч STAT3ЁЂMYD88 Лђ NOD1/2 аХКХЭЈТЗЃЉКЭУтвпЙ§ГЬЃЈШч PD-L1 НЕНтКЭПЙдНЛВцГЪЕнЃЉКмживЊЁЃШЛЖјЃЌзищЕѕЃЛЏЕФживЊадЩаЮДдкЯЕЭГадКьАпРЧДЏЕШздЩэУтвпадМВВЁжаЕУЕНжЄЪЕЁЃ
дкетЯюбаОПжаЃЌЗЂЯж S-зищЕѕЃЛЏЕїНк pDC КЭОоЪЩЯИАћжаЕФ TLR9 ЗДгІЁЃTLR9 дкАћЭтНсЙЙгђжааЏДјжСЩйСНИі S-зищЕѕЃЛЏЮЛЕуЁЃзищЕѕЃЛЏбЛЗЪМгкИпЖћЛљЬхЃЌDHHC3 зищЕѕЃЛЏ TLR9ЃЌНсЪјгкШмУИЬхЃЌЦфжа PPT1 ЭбзищЕѕЃЛЏ TLR9ЁЃTLR9зищЕѕЃЛЏбЛЗжБНгЕїНкХфЬхНсКЯКЭpDCКЭОоЪЩЯИАћЕФЯИАћвђзгЗжУкЁЃвђДЫЃЌдкзЊЛљвђSLEаЁЪѓФЃаЭжаЃЌPPT1вХДЋШБЯнЛђPPT1вжжЦМСПЩНЕЕЭздЩэПЙЬхЫЎЦНВЂМѕЧсЩібзЁЃзищЕѕЃЛЏвжжЦМСЛЙвжжЦЯЕЭГадКьАпРЧДЏЛМепШЫ pDC ВњЩњЕФ IFNІСЁЃбаОПИХЪіСЫвЛжжЕїНкTLR9ЗДгІЕФЗвыКѓаоЪЮЛњжЦЃЌВЂЬсГіСЫжЮСЦздЩэУтвпадМВВЁЕФЧБдкУтвпжЮСЦАаЕуЁЃ
TLR9 КЭ TLR7 ЪЧ S-зищЕѕЃЛЏ
PPT1 ЪЧвЛжжВЮгыШЅзищЕѕЃЛЏЕААзжЪЕФУИЁЃЮЊСЫбаОПзищЕѕЃЛЏдкSLEжаПЩФмЕФзїгУЃЌЮвУЧЗжЮіСЫМИЯюзищЕѕЃЛЏЕААзжЪзщбЇЦзбаОПЁЃTLR9БЛЗЂЯжДцдкгквЛИіЕААзжЪзщЪ§ОнПтжаЁЃвђДЫЃЌЮвУЧПЊЪМЬНЫїTLR9ПЩФмБЛзищЕѕЃЛЏЕФПЩФмадЁЃЪЙгУЕуЛїЛЏбЇПЩвдзМШЗМьВтзищЕѕЃЛЏЕФЫљгаЮЛЕуЃЌАќРЈ S-зищЕѕЃЛЏКЭ O-зищЕѕЃЛЏЮЛЕуЁЃЪЙгУЕўЕЊЛљзищЕЫсЃЈвВГЦЮЊClick-iTзищЕЫсЃЌЕўЕЊЛЏЮяЃЉБъМЧЃЌЮвУЧЗЂЯждкRAW264.7аЁЪѓОоЪЩЯИАћЯЕжаБэДяЕФаЁЪѓTLR9ЃЈmTLR9ЃЉБЛзищЕѕЃЛЏѕЃЛљ-ЩњЮяЫиНЛЛЛ ЃЈABEЃЉ ЪЧСэвЛжжЬивьадМьВт S-зищЕѕЃЛЏЕФДЋЭГЗНЗЈЁЃЮвУЧНјааСЫABEВЂШЗШЯmTLR9ЪЧS-зищЕѕЃЛЏЁЃCalnexin ЪЧвЛжжБэеїСМКУЕФ S-зищЕѕЃЛЏЕААзЃЌгУзїбєадЖдееЃЌRAB7 гУзївѕадЖдееЁЃСНжжЩњЛЏЪдбщОљБэУїаЁЪѓTLR9БЛзищЕѕЃЛЏЁЃ
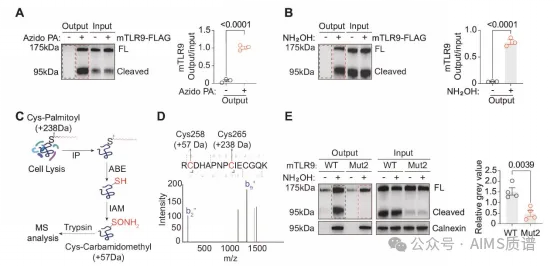
ЛљгкгыABEЯрЫЦЕФдРэЃЌЮвУЧЖдmTLR9НјааСЫжЪЦзЗжЮіЁЃЮвУЧЗЂЯжСНИіАыызАБЫсВаЛљ C258 КЭ C265 БЛ S-зищЕѕЃЛЏЁЃетСНжжАБЛљЫсЮЛгкАћЭтНсЙЙгђжаИЛКЌССАБЫсЕФжиИДађСа 8 ЃЈLRR8ЃЉ КЭ LRR9 жЎМфЕФаЁЛЗжаЁЃШЛЖјЃЌетСНИізищЕѕЃЛЏЮЛЕуВЛдкZЛЗжаЃЌМДЪЕМЪЕФХфЬхНсКЯВлЁЃШЛКѓЃЌЮвУЧМЬајгУБћАБЫсЃЈC258A + C265AЃЌMut2ЃЉШЁДњетСНжжАыызАБЫсЃЌВЂдкTLR9ШБЯнЕФRAW264.7ОоЪЩЯИАћЯЕжаЮШЖЈБэДяИУЭЛБфЬхЁЃS4C-E).дкABEВтЖЈжаЃЌЮвУЧЗЂЯжC258AКЭC265AЭЛБфЪЙmTLR9ЕФзищЕѕЃЛЏЫЎЦННЕЕЭСЫ3БЖЁЃгЩгкTLR7дкSLEЗЂВЁЛњжЦжавВЦ№зХживЊзїгУЃЌЮвУЧбаОПСЫTLR7ЕФзищЕѕЃЛЏзДЬЌЁЃЪЙгУЕуЛїЛЏбЇКЭ ABE ВтЖЈЃЌЮвУЧЗЂЯж mTLR7 ЪЧ S-зищЕѕЃЛЏЁЃзмжЎЃЌЮвУЧЗЂЯжаЁЪѓ TLR9 КЭ TLR7 ЪЧ S-зищЕѕЃЛЏЁЃ
НгЯТРДЃЌМьВщСЫШЫTLR9ЃЈhTLR9ЃЉЕФзищЕѕЃЛЏзДЬЌЁЃABE ВтЖЈБэУїЃЌдкШЫЕЅКЫЯИАћЯЕ THP-1 Лђ 293 T ЯИАћжаБэДяЕФ hTLR9 вВБЛ S-зищЕѕЃЛЏЁЃдкетРяЃЌЮвУЧЗЂЯжTLR9дкШЫРрЯИАћжаБЛS-зищЕѕЃЛЏЁЃ
НсТл
НсЙћБэУїЃЌTLR9зищЕѕЃЛЏбЛЗгаСНИіжївЊВНжшЃКИпЖћЛљЬхжаЕФзищЕѕЃЛЏКЭФкЬхжаЕФШЅзищЕѕЃЛЏЁЃЭЈЙ§НјааЯИАћЗжРыЪЕбщЃЌЮвУЧЗЂЯжDHHC3ШБЯнЯИАћдкИпЖћЛљЬхжаБЃСєСЫИќЖрЕФШЋГЄTLR9ЃЌЖјдкФкЬхжаЗЂЯжСЫНЯЩйЕФСбНтTLR9ЁЃвђДЫЃЌзищЕѕЃЛЏЪЧНЋTLR9ДгИпЖћЛљЬхзЊдЫЕНФкЬхЫљБиашЕФЁЃФкЬхжаНЯЩйСбНтЕФ TLR9 НЋЕМжТ TNF ЗжУкМѕЩйЃЌетдк DHHC3 ШБЯнЯИАћжаЙлВьЕНЁЃЙигкШЅзищЕѕЃЛЏВНжшЃЌЪЙгУCo-IPЪЕбщЬНВтUNC93B1ЩЯБЃСєЕФTLR9ЕФСПЃЌЮвУЧЗЂЯждкHDSFДІРэЕФЯИАћжагыUNC93B1НсКЯЕФTLR9НЯЩйЃЌетвВЕМжТTLR9аХКХДЋЕММѕЩйЁЃзмжЎЃЌTLR9 зищЕѕЃЛЏЕїНк TLR9 ДгИпЖћЛљЬхЕНФкЬхЕФдЫЪфЃЌTLR9 ШЅзищЕѕЃЛЏПижЦФкЬхжаUNC93B1ЪЭЗХ TLR9ЁЃвжжЦШЮвЛВНжшЖМЛсЕМжТФкЬхжагЮРыСбНтЕФTLR9МѕЩйЃЌДгЖјМѕЩйаХКХДЋЕМЁЃгыДЫвЛжТЃЌHDSFжЮСЦКЭPPT1ШБЗІОљЯджјМѕЛКСЫSLEЕФЗЂВЁЛњжЦЁЃШЛЖјЃЌ2-BPДІРэЖдЬхФкздЩэПЙЬхЕФВњЩњгАЯьВЛДѓЁЃЖд DHHC3 ШБЯнаЁЪѓЕФНјвЛВНбаОПгІНјвЛВНВћУї TLR7/9 зищЕѕЃЛЏЕФживЊадЁЃ
TLR7 вбБЛжЄУїЪЧ SLE ЗЂВЁЛњжЦЕФживЊаХКХЭЈТЗЁЃЪЙгУЕуЛїЛЏбЇКЭ ABE ВтЖЈЃЌЮвУЧЗЂЯж TLR7 вВЪЧ S-зищЕѕЃЛЏЁЃШЛЖјЃЌDHHC3 Й§БэДяВЛФмЪЙ TLR7 зищЕѕЃЛЏЃЌетБэУїЦфЫћ DHHC ПЩФмЪЧ TLR7 зищЕѕЃЛЏЕФдвђЁЃИќживЊЕФЪЧЃЌABEЗжЮіЯдЪОЃЌTLR7дкR848МЄЛюКѓЮДБЛШЅзищЕѕЃЛЏЁЃЯрБШжЎЯТЃЌTLR9 дк CpG МЄЛюКѓбИЫйЭбзищЕѕЃЛЏЃЌМДЪЙдк 24 аЁЪБКѓШдБЃГжЭбзищЕѕЃЛЏЁЃДЫЭтЃЌPPT1 ЕФЙ§БэДяВЛЛсЕМжТ TLR7 ШЅзищЕЫсЛЏЁЃPPT1вжжЦМСHDSFвВВЛФмИФБфTLR7зищЕѕЃЛЏЫЎЦНЁЃзмжЎЃЌTLR7 ВЛВЮгыгы TLR9 ЯрЭЌЕФзищЕѕЃЛЏ/ШЅзищЕѕЃЛЏбЛЗЁЃСбНтЕФ TLR7 ВЛашвЊДгUNC93B1жаЪЭЗХЃЌЖј TLR9 ашвЊЁЃPPT1 Жд TLR9 ЕФШЅзищЕѕЃЛЏДйНјСЫЦфдкШмУИЬхжаХфЬхНсКЯжЎЧАДгUNC93B1ЪЭЗХЃЌЖј TLR7 ПЩФмВЛашвЊШЅзищЕѕЃЛЏРДЦєЖЏЯТгЮаХКХДЋЕМЁЃвђДЫЃЌдк PPT1 ШБЯнаЁЪѓжаЙлВьЕНЕФБэаЭВЛЬЋПЩФмжївЊЪЧгЩгк TLR7 ЕФЮЩТвШЅзищЕѕЃЛЏЁЃ
Ъ§ОнНвЪОСЫвЛжжЕїНкаЁЪѓКЭШЫРр TLR9 КЭ TLR7 ЗДгІЕФИпЖћЛљЬхШмУИЬхзищЕѕЃЛЏЭООЖЁЃЮвУЧЗЂЯжЃЌетжжФкЬхзищЕѕЃЛЏ/ШЅзищЕѕЃЛЏбЛЗЕФЦЦЛЕвжжЦСЫ pDC КЭОоЪЩЯИАћжа TLR9 ХфЬхНсКЯКЭЯИАћвђзгЕФВњЩњЁЃPPT1 вХДЋШБЯнЛђЛЏбЇвжжЦвжжЦСЫ B6 жаЕФПЙ DNA здЩэПЙЬхВЂМѕЧсСЫЩібзЁЃSle1yaaаЁЪѓЁЃМјгкЦфдк cDC1 КЭ pDC жаЕФЯрЗДзїгУЃЌPPT1 ПЩФмЪЧ SLE жЮСЦЕФЧБдквЉЮявжжЦАаЕуЃЌвђЮЊ PPT1 ПЩФмОпгадіЧП CTL ЖдАЉжЂКЭВЁдЬхЗДгІЕФЖюЭтвцДІЁЃ
БОЮФФПЕФЕААзTLRЗжзгзищЕѕЃаоЪЮжЪЦзМьВтЪдМСКагЩAimsmass Co.LtdЬсЙЉЃЌШчЩЯЭМЫљЪОЃЌЖдmTLR9НјааСЫжЪЦзЗжЮіЁЃЮвУЧЗЂЯжСНИіАыызАБЫсВаЛљ C258 КЭ C265 БЛ S-зищЕѕЃЛЏЁЃетСНжжАБЛљЫсЮЛгкАћЭтНсЙЙгђжаИЛКЌССАБЫсЕФжиИДађСа 8 ЃЈLRR8ЃЉ КЭ LRR9 жЎМфЕФаЁЛЗжаЁЃ
еыЖдгкзищЕѕЃаоЪЮЃЌНќМИФъЪБМфЗЂБэЕФИпЗжЮФЯзЃЌОљЪЧЙигкЬиЖЈФПБъЕААзаоЪЮЕФбаОПЃЌвдМАЩЯЯТгЮЯрЙизїгУЕААзЗжзгЕФНЛЛЅЫљв§Ц№ЕФЩњЮябЇаЇгІЃЌзищЕѕЃаоЪЮБОЩэЪЧЯрЖдгкГЃЙцСзЫсЛЏЃЌЗКЫиЛЏвдМАввѕЃЛЏЕШаоЪЮЗЂЩњНЯЩйЕФвЛжжжЌѕЃаоЪЮЃЌБОЮФПЦбЇМвЭЈЙ§ЕїбаЧАЦкЦфЫћПЦбаЙЄзїепЕФзщбЇЪ§ОнЭЈЙ§НјааПьЫйЕФWBМьВтЃЈПЩПМТЧAimsmassЪдМСКаЃЌAM10313&AM10314ЃЉЃЌЫцКѓЭЈЙ§жЪЦзMSМьВтЦфзищЕѕЃаоЪЮЮЛЕуЃЌЕУЕНПЩППзищЕѕЃаоЪЮЪ§ОнЁЃ
Mass spectrometry
Sample preparation with ABE was previously described68,69. RAW264.7 cells carrying mTLR9-Flag were lysed, solubilized and immunoprecipitated with anti-FLAG affinity beads (Sigma, A2220). After washing with 1 ml of lysis buffer three times at 4 ЁуC, the beads were subjected to the ABE assay preparation procedure and an alkylation reaction using an IP-ABE Palmitoylation Kit for MS (Aims, AM10417) following the protocols on the manufacturerЁЏs instruction sheet. The eluted sample was used for further MS analysis.
зЊдиГіДІЁОNature Communication IF=17ЁПжмЦкадзищЕѕЃЛЏЕїНкаЁЪѓЕФ TLR9 аХКХДЋЕМКЭздЩэУтвп
дЮФГіДІЃКhttps://www.nature.com/articles/s41467-023-43650-z
зищЕѕЃаоЪЮhttps://www.aimsmass.com/s-palmitoylation.html
ЬиЖЈЕААззищЕѕЃаоЪЮУтвпгЁМЃЪдМСКа https://www.aimsmass.com/products/AM10313.html
ЬиЖЈЕААззищЕѕЃаоЪЮУтвпгЁМЃЪдМСКаЃЈКЌHRPЃЉhttps://www.aimsmass.com/products/AM10314.html
ЬиЖЈЕААззищЕѕЃаоЪЮжЪЦзЪдМСКа https://www.aimsmass.com/products/AM10417.html
https://wap.sciencenet.cn/blog-3402731-1486853.html
ЩЯвЛЦЊЃК[зЊди]ЁОIF=21 Nature MetabolismЁПвьЮьЯЉзщбЇЗўЮёЙњФкЪзЗЂ ЗЈФсЛљаоЪЮЭЈЙ§ЯпСЃЬхдіЧПздЩэУт..
ШЋВПзїепЕФЦфЫћзюаТВЉЮФ
ШЋВПОЋбЁВЉЮФЕМЖС
- • ЛљН№БОзгЁАЪнЩэЬсжЪЁБЃЌХфЭМВЛвЊВШет3жжПгЃЁ
- • зЃКиЃЁПЦбЇЭј2025ФъЖШЪЎМбВЉЮФЦРбЁЛюЖЏНсЙћНвЯў
- • ЮїББЙЄвЕДѓбЇеХЧягэ&еХКЭХє&ЭѕЬьЫЇЕШЃКИФадВЛПЩФцМќCOFsЪЕЯжИпаЇCO₂ЛЙд
- • ББОЉЪаПЦбЇММЪѕЮЏдБЛсЁЂжаЙиДхПЦММдАЧјЙмРэЮЏдБЛсЙњМЪКЯзїДІМАЙњМЪжааФвЛааЕНЗУMDPIББОЉЪаЭЈжнАьЙЋЪв
- • КЃбѓжаЕФАЕбѕ
- • ЩЯЮчЭЖЯТЮчUnder ReviewЃЌОоКУЭЖЕФЫФЧјSCIРДИхОЭЪеЃЌ0АцУцЗбЃЁ