
七、三羧酸循环:在线粒体基质中的彻底氧化与碳骨架重组
三羧酸循环是细胞呼吸的核心枢纽,其本质是在线粒体基质中,完成对燃料分子的彻底氧化与碳骨架重组。它是生物能学中,将物质分解与能量提取推向极致的分子级“精密熔炉”。
为了直观把握这一复杂过程,图5可以清晰地展示循环的完整路径、核心输入与关键产出:
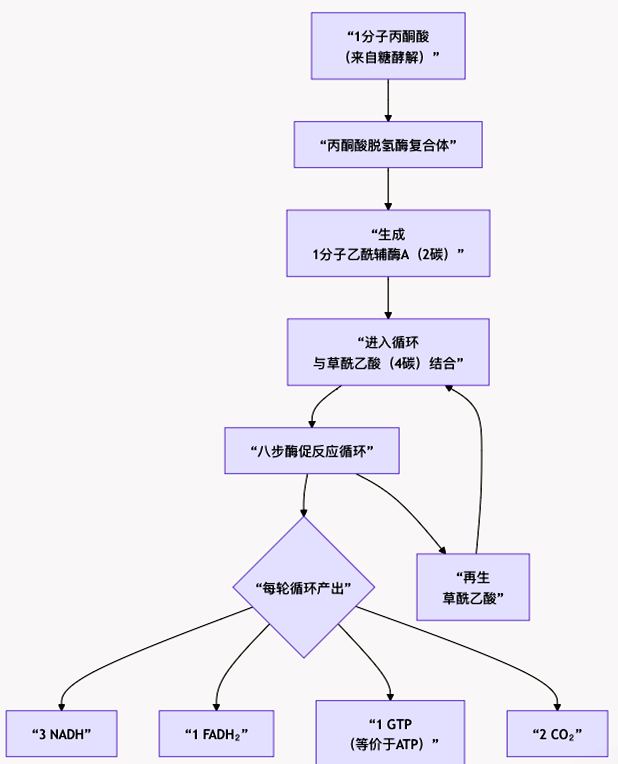
图5 三羧酸循环的完整路径、核心输入与关键产出
下面,我们来深入解读这个循环如何实现“彻底氧化”与“碳骨架重组”。
1. “彻底氧化”:燃烧至分子骨架的终极拆解
循环的氧化是彻底的,其目标是将乙酰基(-COCH₃)中的碳原子完全转化为CO₂,并榨取出所有可用的高能电子。
(1)起点:来自糖、脂肪、蛋白质降解的乙酰辅酶A(2碳单位)进入循环。
(2)过程:在八步反应中,通过四次脱氢反应(产生3个NADH和1个FADH₂)和两次脱羧反应(释放2个CO₂),乙酰基被完全拆解。
(3)结果:进入循环的2个碳原子,最终以2分子CO₂的形式被“呼出”。更重要的是,碳原子中蕴藏的化学能,几乎全部以高能电子的形式,储存于NADH和FADH₂中,它们将进入呼吸链,驱动合成大量ATP。
2. “碳骨架重组”:循环再生的动态平衡
循环的精妙之处在于,它在分解乙酰基的同时,必须再生出起始受体——草酰乙酸,以维持循环持续运转。这八步反应,本质上是一个4碳骨架(草酰乙酸)不断结合2碳单位(乙酰基)、经过一系列重排、氧化、最后又变回4碳骨架的过程。碳原子在循环中不断改变连接方式(重组),但载体的数量(碳骨架长度)在循环结束时恢复如初。
3. 能量与物质的精确账目
以1分子葡萄糖(产生2分子丙酮酸)计算,经过三羧酸循环的总账目如下:
表3 经过三羧酸循环的能量与物质总账目
项目 | 数量(每分子葡萄糖) | 去向与意义 |
乙酰辅酶A | 2分子 | 进入循环的“燃料”。 |
CO₂释放 | 4分子 | 碳被彻底氧化的终产物。 |
NADH产生 | 6分子 | 携带大量高能电子,是呼吸链生产ATP的主要驱动力。 |
FADH₂产生 | 2分子 | 携带高能电子,进入呼吸链。 |
GTP(ATP)产生 | 2分子 | 通过底物水平磷酸化直接生成,可立即使用。 |
4. 循环的枢纽地位:代谢网络的十字路口
三羧酸循环远不止是糖代谢的终点,它更是细胞代谢的中央整合平台:
(1)糖代谢:如上所述,是糖酵解的后续。
(2)脂肪代谢:脂肪酸β-氧化产生的大量乙酰辅酶A,也汇入此循环。
(3)蛋白质代谢:多种氨基酸脱氨后的碳骨架,可转化为循环的中间产物(如α-酮戊二酸、草酰乙酸)进入氧化,或用于合成葡萄糖(糖异生)。
5. 思想史意义:机械论解释的又一丰碑
在思想史脉络中,三羧酸循环(由克雷布斯等人于1937年阐明)的发现,是继糖酵解之后,对“生命是化学反应总和”这一机械论纲领的又一次决定性证实。
(1)它展示了极致的“还原”:将生命最核心的呼吸作用,还原为一个在试管中可模拟的、由八种酶精确催化的化学循环。每一步反应、每一个中间产物都清晰可知。
(2)它统一了代谢的“混乱”:它揭示了糖、脂肪、蛋白质这三类看似不同的物质,最终在分子层面汇聚于同一条化学通路被统一处理。这极大地强化了生命的物质统一性观念。
(3)它给活力论留下更小的空间:当这个负责“彻底氧化”、为生命提供90%以上能量的核心过程,被完全描绘为一张化学反应图时,“生命力”在能量代谢领域的解释必要性就彻底归零了。
总而言之,三羧酸循环是生命能量经济的“中央处理器”和“终极氧化熔炉”。它完美体现了现代生物能学的核心思想:生命的神奇,不在于超自然的活力,而在于这些基于物理化学定律演化而来的、极度复杂且精妙的分子机器与反应网络。
如果需要进一步了解这个循环中的某个关键反应(如柠檬酸合酶的作用)、其精细的调控机制,或电子传递链如何具体衔接,我们可以进一步深入。
八、电子传递链与氧化磷酸化:能量转换的最终舞台
电子传递链与氧化磷酸化,是生物能量转换的终极阶段。在这里,之前所有代谢途径(糖酵解、三羧酸循环)中收集的“能量票据”(NADH、FADH₂),将在此“兑现”为巨量的通用货币——ATP。这个位于线粒体内膜上的过程,是现代生物能学的巅峰之作,也是彻底埋葬“活力论”在能量领域解释权的决定性证据。
为了清晰地展示这个复杂系统的全貌,图6整合了从电子传递到ATP合成的完整流程与核心机制:
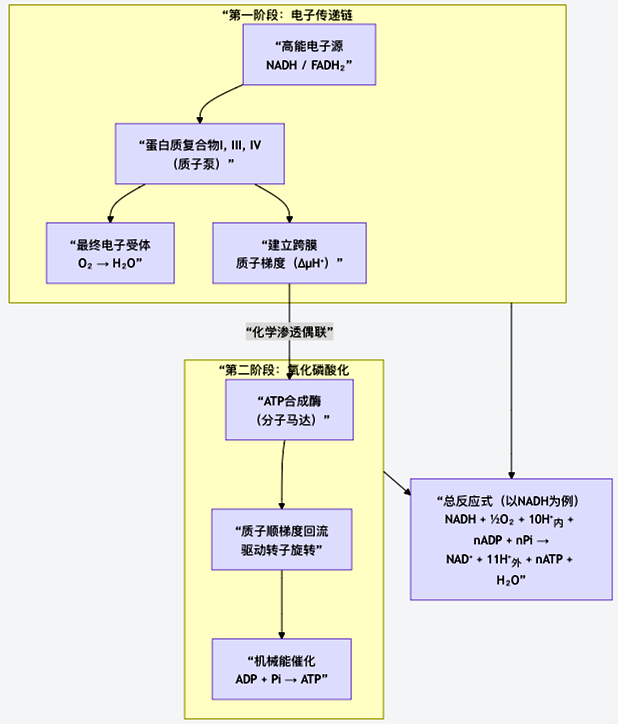
图6 从电子传递到ATP合成的流程与机制
1. 电子传递链——燃烧电子,泵送质子
电子传递链是一系列嵌入线粒体内膜的蛋白质复合物(I、II、III、IV)和移动载体(辅酶Q、细胞色素c)。
(1)电子来源:NADH和FADH₂在此处被氧化,脱下高能电子(e⁻)和质子(H⁺)。
(2)电子“瀑布”:电子像水流跌落瀑布一样,沿着这些复合物从高能级向低能级逐级传递。每一步释放的能量,都用于一个核心任务:将基质中的质子(H⁺)泵入内膜外的膜间隙。
(3)最终归宿:传递链的终端是细胞色素c氧化酶(复合物IV),它将电子、质子和环境中的氧气(O₂)结合,生成水(H₂O)。这是生命需要氧气的分子根源。
(4)核心成果:通过泵送质子,在膜两侧建立起一个强大的电化学梯度(即质子动力势,ΔμH⁺),包括浓度差(化学梯度)和电荷差(电位梯度)。这就像一个蓄满水的水库,蕴藏着巨大的势能。
2. 氧化磷酸化——质子回流,驱动“分子水车”合成ATP
储存的质子势能如何转化为ATP的化学能?答案在于一个精妙的分子机器——ATP合成酶。
(1)质子通道与分子马达:ATP合成酶也嵌在内膜上。当膜间隙高浓度的质子通过它的通道顺梯度流回基质时,就像水流推动水车,驱动了酶内部一个称为“转子”的组件旋转。
(2)旋转催化:转子的机械旋转,引起酶活性中心构象的周期性变化(“结合”“催化”“释放”三种状态),从而将ADP和无机磷酸(Pi)合成为ATP。这是“结合变构机制”的完美体现。
(3)严格偶联:这个过程与电子传递紧密偶联。没有电子传递建立梯度,ATP合成酶就无法工作;同时,质子回流也解除了对电子传递的抑制,使两者速率完美匹配。
3. 能量账目:惊人的产出效率
这是产能效率最高的阶段。理论上,1分子NADH通过此过程可产生约2.5个ATP,1分子FADH₂可产生约1.5个ATP。加上糖酵解和三羧酸循环中直接产生的少量ATP,1分子葡萄糖彻底氧化最多可产生约30-32个ATP,其中超过90%产自这个“最终舞台”。
4. 思想史意义:机械论解释的终极胜利
这一机制的阐明(尤其是彼得·米切尔1961年提出的化学渗透理论),具有划时代的意义:
(1)完成了最关键的“还原”:它将生命最核心、最神秘的能量创造活动(呼吸),还原为一个基于物理(质子梯度、电势差、旋转机械力)和化学(氧化还原、酶催化)原理的清晰模型。酶的“分子经纪人”角色在此达到极致。
(2)提供了统一的理论:它统一解释了线粒体、叶绿体和细菌的能量转换,揭示了生命世界在分子层面的深层次统一性。
(3)彻底消解了“活力”的空间:当科学家能在人工脂质膜上重建质子梯度并驱动ATP合成时,当ATP合成酶的原子分辨率结构图清晰地展示出这个“分子水车”时,任何关于能量转换需要“特殊生命力”的假设都变得完全多余且不可想象。
(4)结论:电子传递链与氧化磷酸化,是生命将物质燃烧(氧化)与能量货币(ATP)铸造进行精密耦合的终极解决方案。它不仅是生物能学的“最终舞台”,也是科学对活力论取得完全、彻底胜利的最终舞台,标志着生命科学完全确立了其物理化学基础。
如果我们希望了解化学渗透理论提出时的科学争议,或ATP合成酶更精妙的结构工作机制,可以继续深入。
九、光合作用如何捕获和转换光能?
(未完待续)
转载本文请联系原作者获取授权,同时请注明本文来自聂广科学网博客。
链接地址:https://wap.sciencenet.cn/blog-279293-1514725.html?mobile=1
收藏