博文
生物入侵之遗传悖论
 精选
精选
||
奠基者效应(Founder effects):一个大种群中少部分个体迁移或传播至另一区域,且与原来的种群相隔离。经一段时间的繁衍,种群个体数虽然增加,但整个族群的遗传多样性却未明显提升。

阿利效应(Allee effects):当种群大小或密度在一定范围时,种群的适合度于种群大小或密度之间存在正相关的现象。若种群数量或密度过小时,会有灭绝的危险。

在一种外来物种刚入侵到新的生境时,往往只涉及祖先物种里的少数群体或个体,从而导致形成的新入侵群体遗传多样性很低,其种群的适合度也较低。尽管瓶颈效应使得新入侵的群体遗传多样性很低,且种群的适合度也较低,但这些新入侵的外来物种群体仍能够适应新生境,并能够通过适应性进化(Adaptive evolution)形成星火燎原之势。这种很低的遗传多样性和很强的适应能力之间的巨大反差被称为“生物入侵的遗传悖论”(Genetic paradox of biological invasion)。
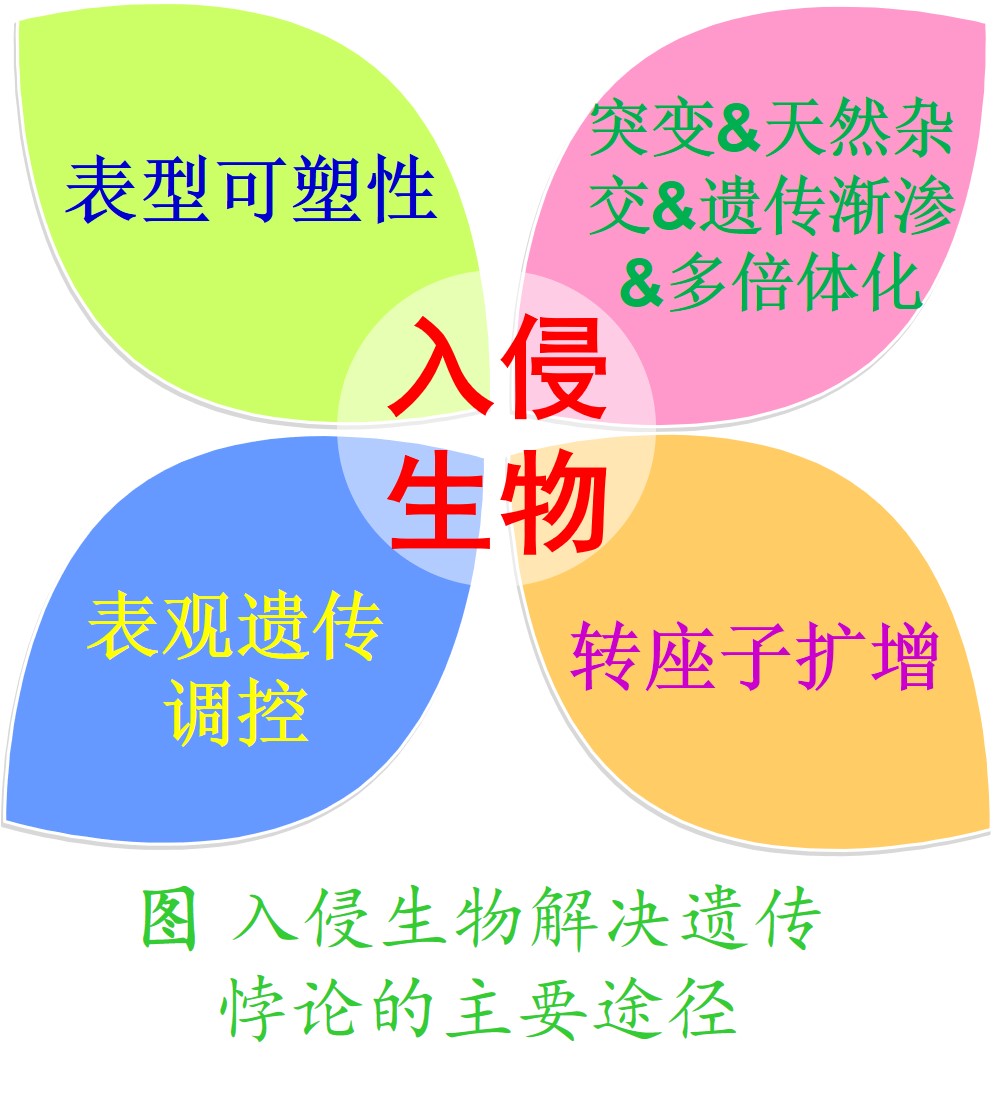
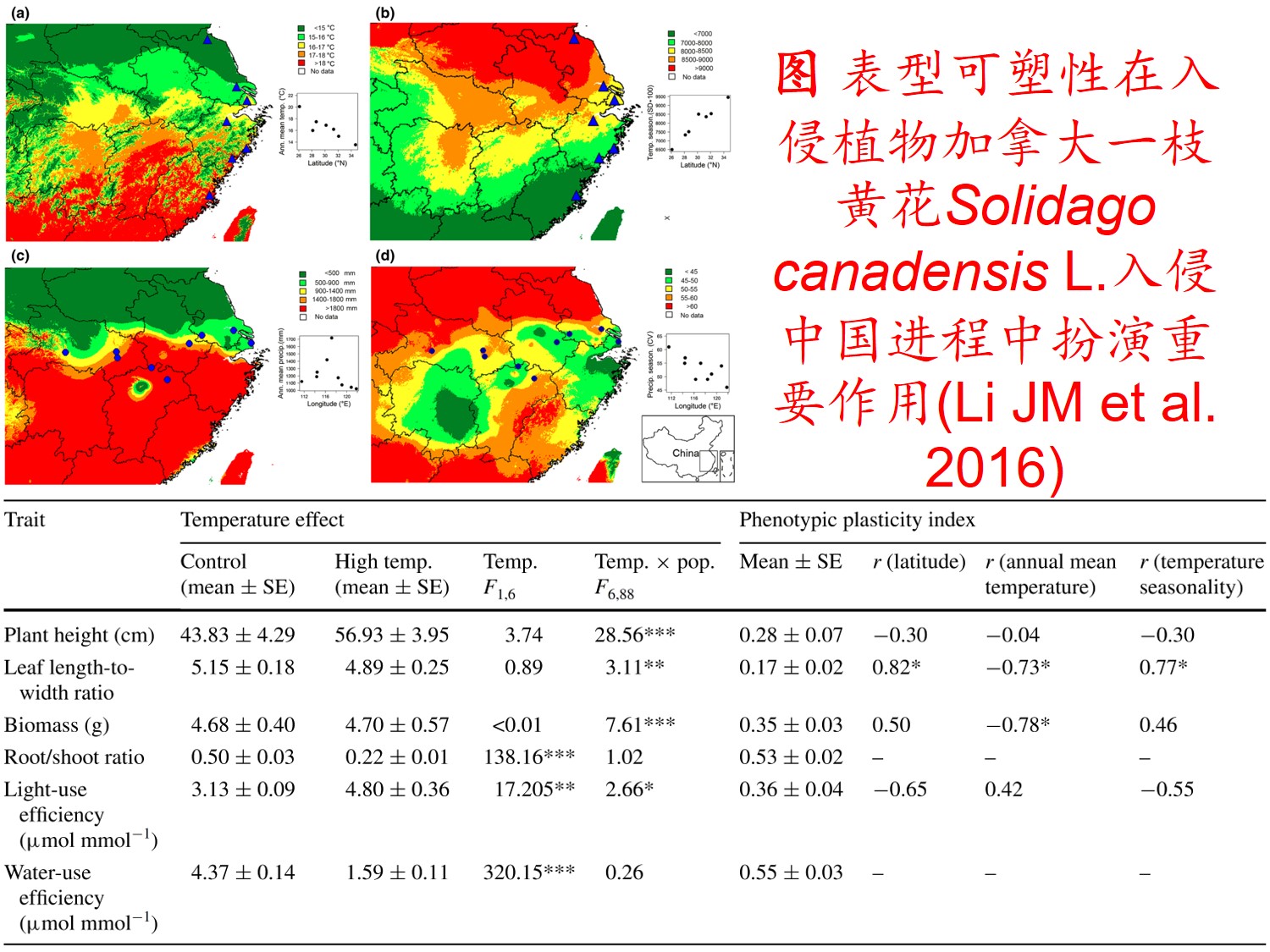
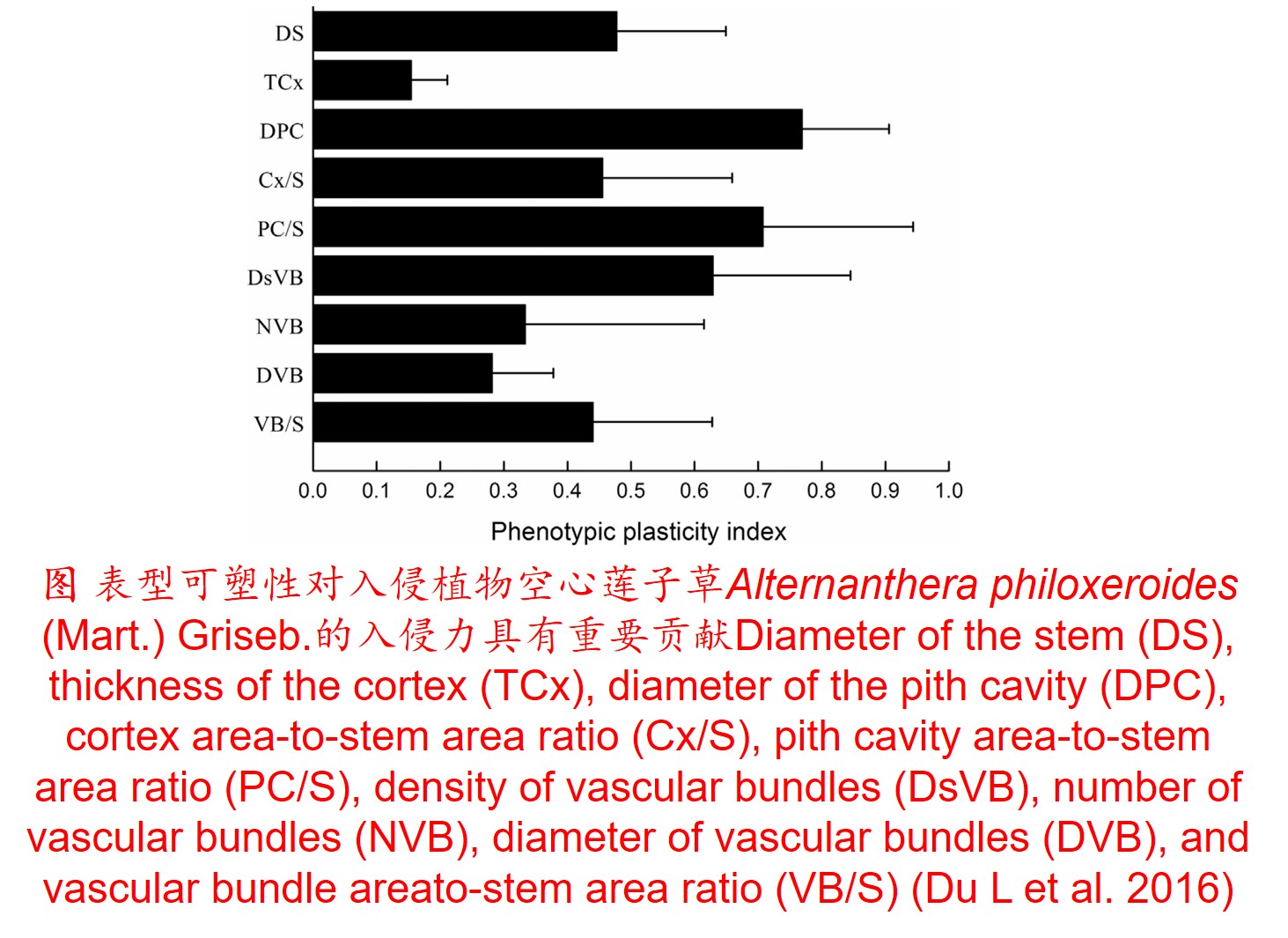
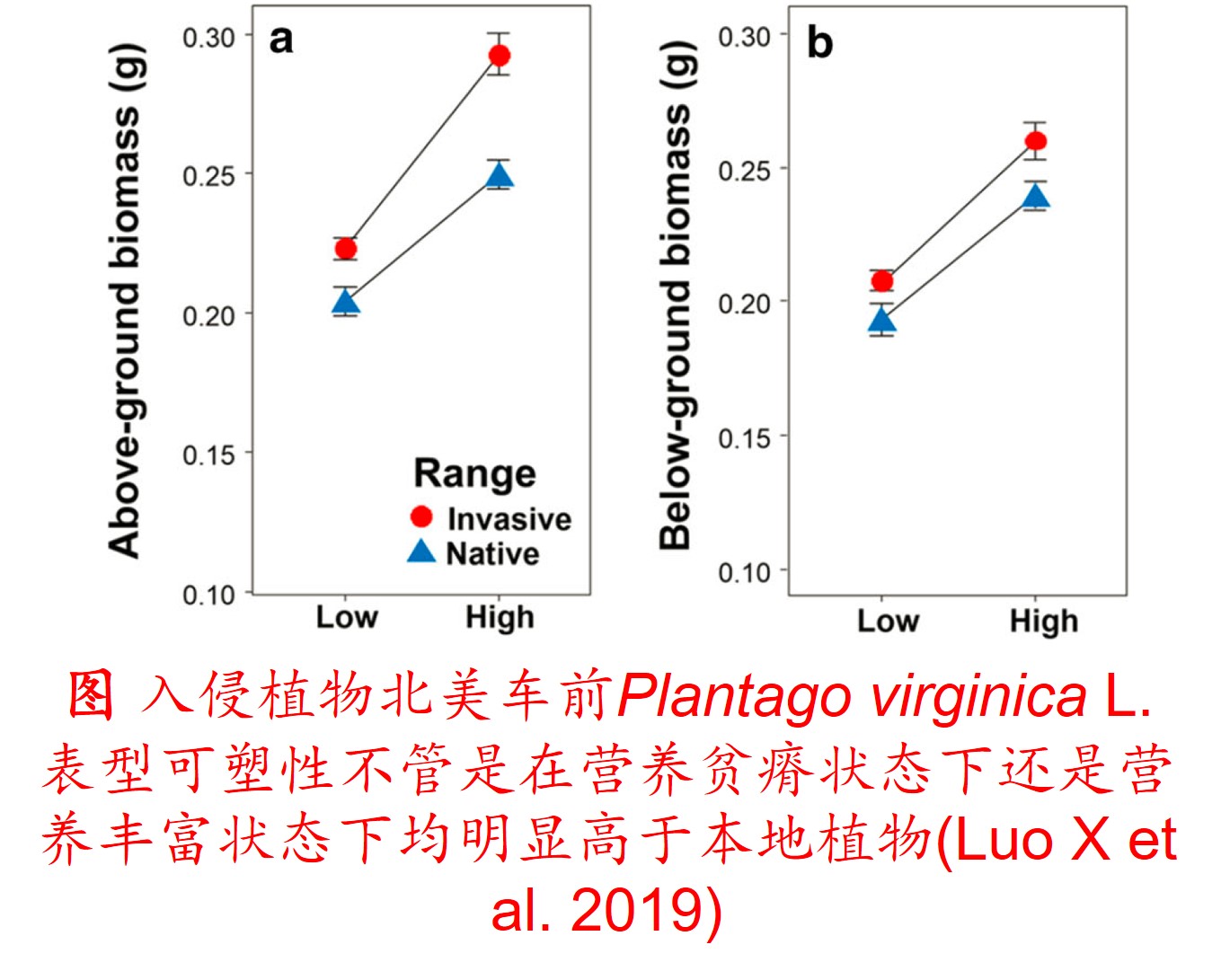
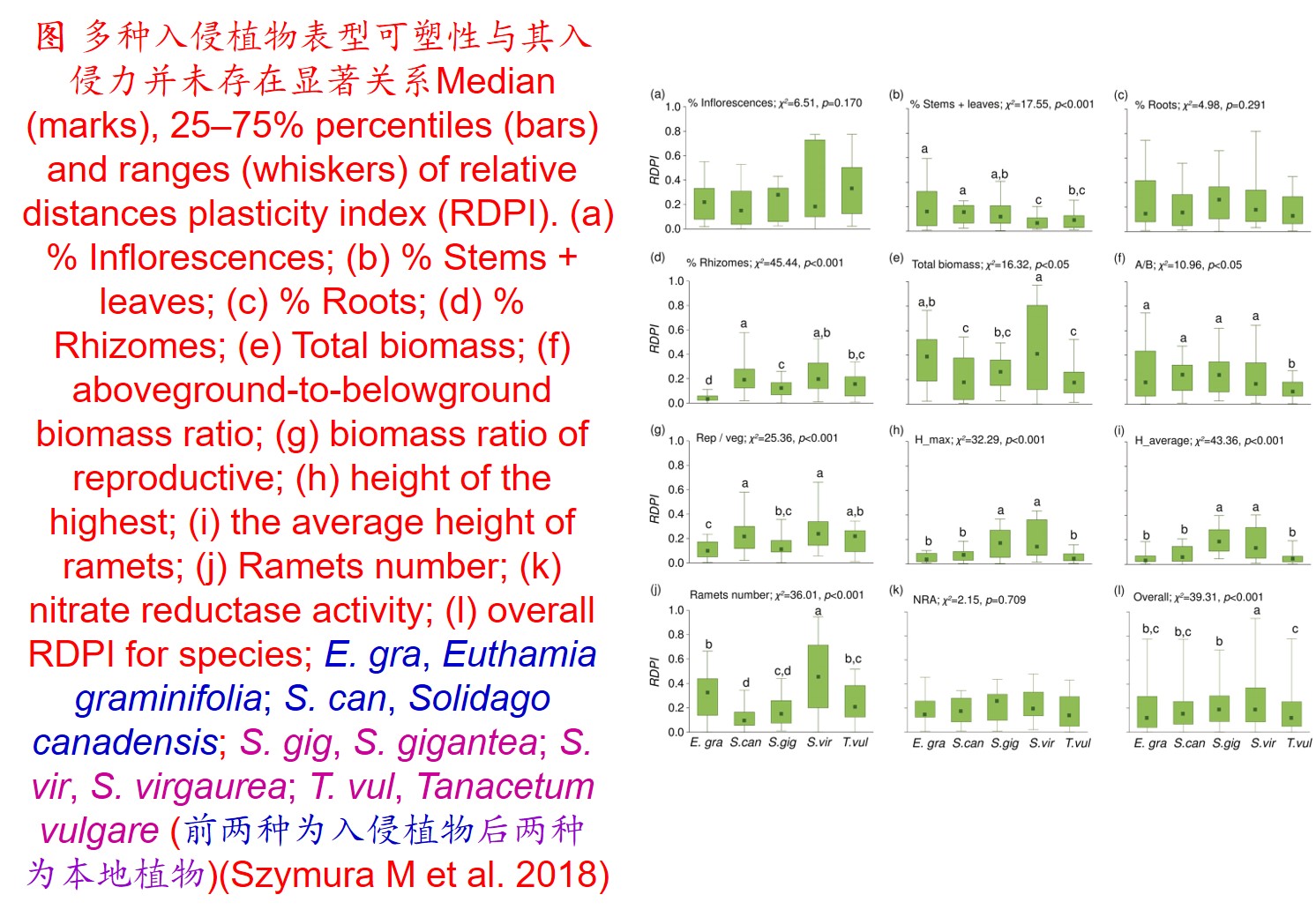
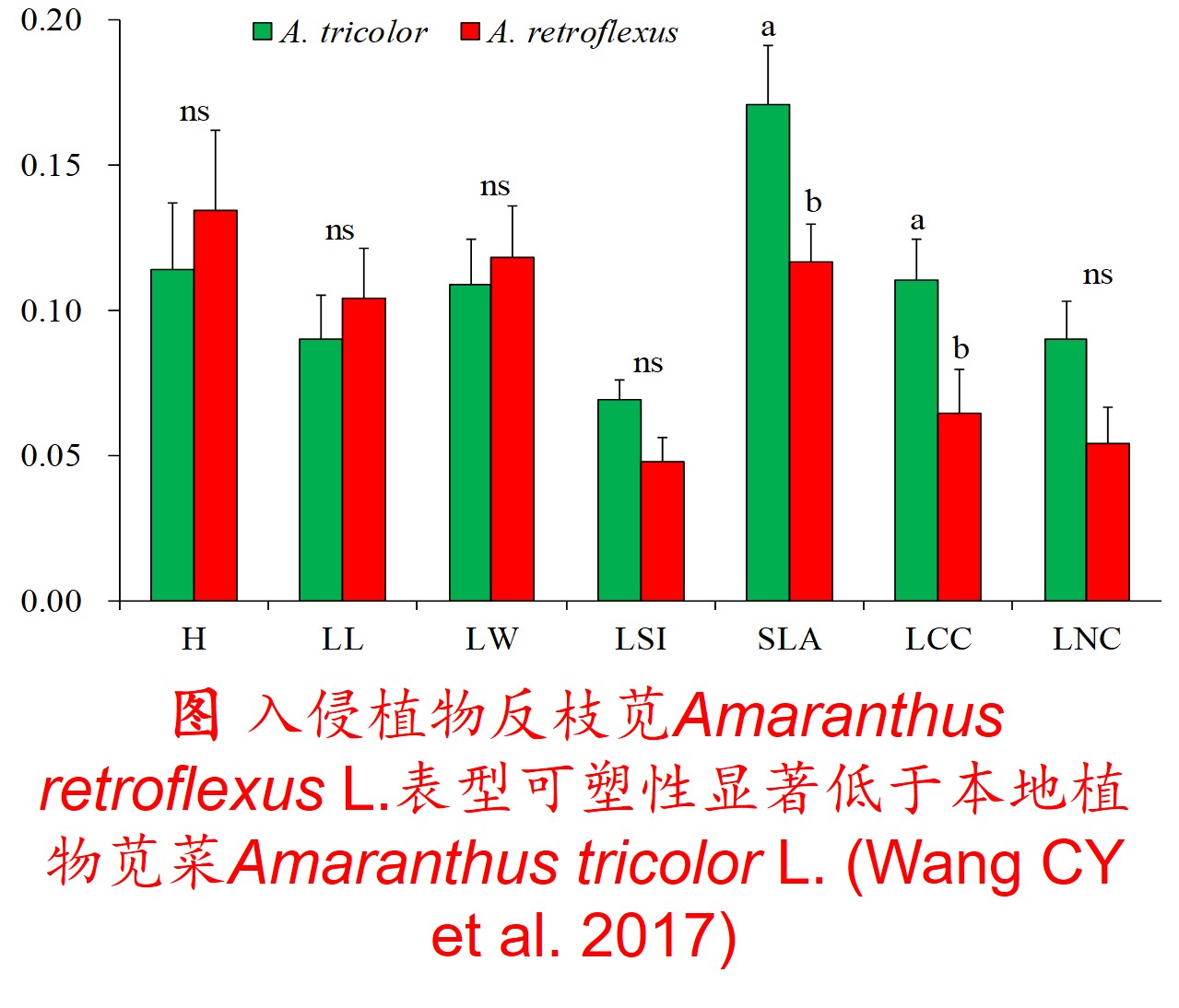
1. 表型可塑性(Phenotypic plasticity):
在环境影响下,某一基因型在表型上产生变异的能力,包括形态、生理、生长、代谢和生活史等方面的塑性。

(1)植物变异系数(The coefficient of variation, CV) (Bhoj D & Ahsanullah M 1993; Forkman J 2009; Krishnamoorthy K & Lee M 2014):
𝐶𝑉=𝜎/𝜇
式中μ为调查样方内各物种功能性状的平均值,σ为调查样方内各物种功能性状的标准差(Standard Deviation)。CV值越大植物功能性状变异性越高。
(2)植物表型可塑性指数(Phenotypic plasticity index, PI) (Funk JL 2008; Chen LY et al. 2013; Lamarque LJ et al. 2013):
𝑃𝐼=(𝑀𝑎𝑥−𝑀𝑖𝑛)/𝑀𝑎𝑥
式中Max为同一处理下同一植物生理生态特性指标的最大值,Min为同一处理下同一植物生理生态特性指标的最小值。

2. 突变(Mutation):
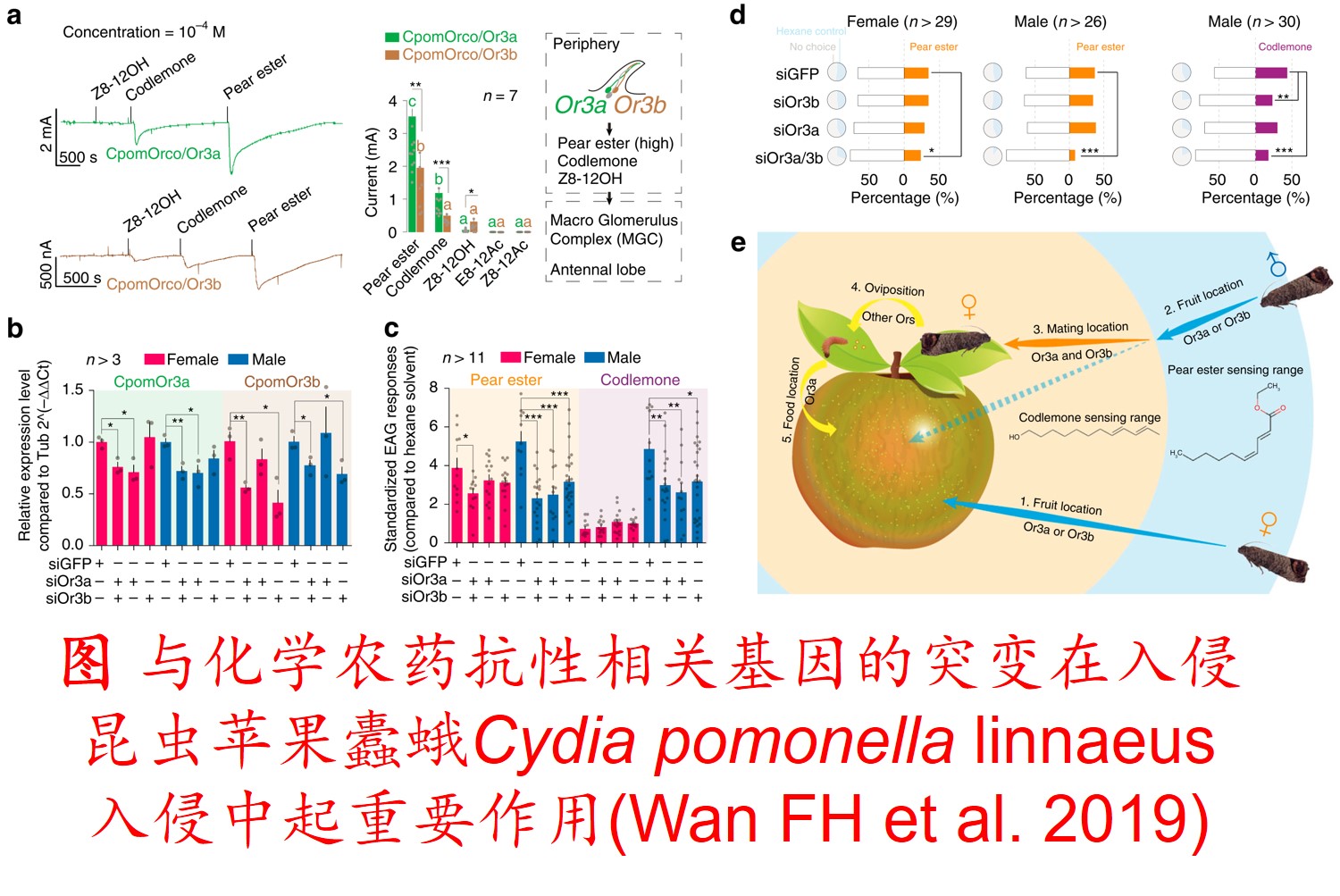
突变一般是指生物体DNA基因组核苷酸序列的改变。突变主要是由于DNA复制(特别是减数分裂)出错或DNA损伤(如暴露于辐射或致毒物等引起)后错误的修复造成的。一般情况下,突变可将新等位基因引入生物群体并进而增加群体的遗传变异,从而增强其适应性进化。此外,一些基因的突变并未导致其编码蛋白质的氨基酸序列的变化,这样的突变被称为沉默突变(Silent mutations)。
3. 天然杂交(Natural hybridization)和遗传渐渗(Genetic Introgression):
天然杂交是植物有性生殖的重要过程,在其进化和物种形成的过程中具有重要作用。通常杂交是指遗传上具有差异的群体或具有不同基因型的个体间进行有性生殖的过程。植物通过不同个体间的传粉和受精来实现。杂交可发生在同一物种的不同群体或同一群体的个体之间,即种内杂交(Intraspecific crossing),也可发生在不同物种的个体之间,即种间杂交(Interspecies mating) (Rieseberg & Carney 1998; Ellstrand & Schierenbeck 2000)。

其中种内杂交是自然界中最常见的现象之一,即使是最严格的自花授粉植物(如大豆和大麦),也会有一定频率的杂交事件发生。种间杂交发生的概率相对较低,在植物中发生的概率约25% (Mallet 2005)。



狭义的遗传渐渗是指基因或遗传物质通过群体中的杂种个体与其亲本个体之间的不断回交而导致基因在群体或个体之间转移和传递的过程,它是物种形成和适应性进化的一个非常重要的遗传机制。而广义的遗传渐渗是指基因或遗传物质在有一定遗传差异的个体或群体之间进行转移和传递的过程(Rieseberg & Carney 1998)。
通过自然杂交和遗传渐渗这一连续的过程,杂种与其亲本将会在个体的遗传基础和群体的遗传多样性水平上发生明显的变化,进而影响其适合度(Baack & Rieseberg 2009)。
相关研究表明:天然杂交和遗传渐渗可改变入侵植物对环境的适应性并明显提高其生长竞争力。特别是对于转基因植物的外源基因是否会通过天然杂交(基因漂移)和遗传渐渗而导入野生近缘种,并通过改变野生近缘种的适合度和生存竞争力,而使其可能变为入侵物种。

如杂草稻可能是野生稻与栽培稻经自然杂交和遗传渐渗产生。更重要的是杂草稻在一些地区已完全具备入侵物种的关键生理生态特性,并已造成了显著的生态危害。




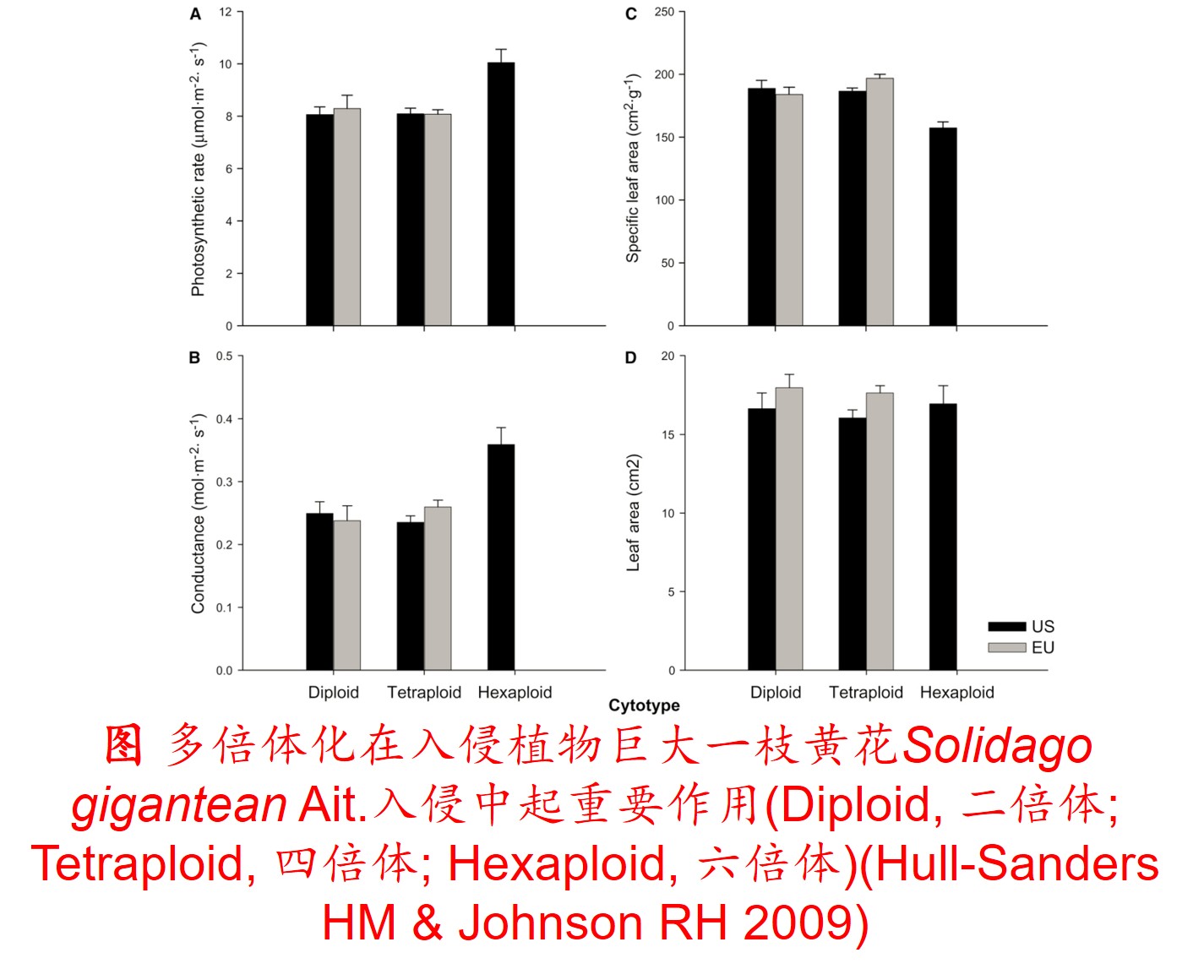
4. 多倍体化(Polyploidization):
植物的天然杂交和染色体加倍是导致多倍体物种产生的主因。多倍体化在高等生物进化中发挥重要作用,多倍化是生物演化的内在动力,它们给予生物体基因组的与功能的可塑性,特别是多倍化驱动空间生态位分化,并显著提高种群耐胁迫能力、遗传多样性和适合度。
也有学者认为多倍体代表基因组更能忍受变化的过渡阶段,多倍体化是细胞应对压力的一种自我保护机制。
多倍体包括同源多倍体(Autopolyploid)和异源多倍体(Allopolyploid)。同源多倍体是指包含两个或两个以上具有相同来源基因组的多倍体,通常是由同一物种的未减数配子产生融合,或正常减数配子之间的杂交和染色体加倍而产生;而异源多倍体则是指包含两个或两个以上具有不同来源基因组的多倍体,往往是由不同物种之间的个体经天然杂交和染色体加倍而产生的。


多倍体的形成可对物种或群体的适合度产生较大影响。由于多倍体植物含双亲的基因组,比其二倍体亲本物种具有成倍增加的染色体数目,且在多倍体形成的过程中经历复杂的遗传重组,故大多数多倍体植物不仅体型较大,且比其亲本种具有更强的适应性和竞争能力(Feldman & Levy 2009)。所以,多倍体植物成为入侵物种的可能性显著高于其二倍体亲本(Prentis PJ et al. 2008; Feldman & Levy 2009)。






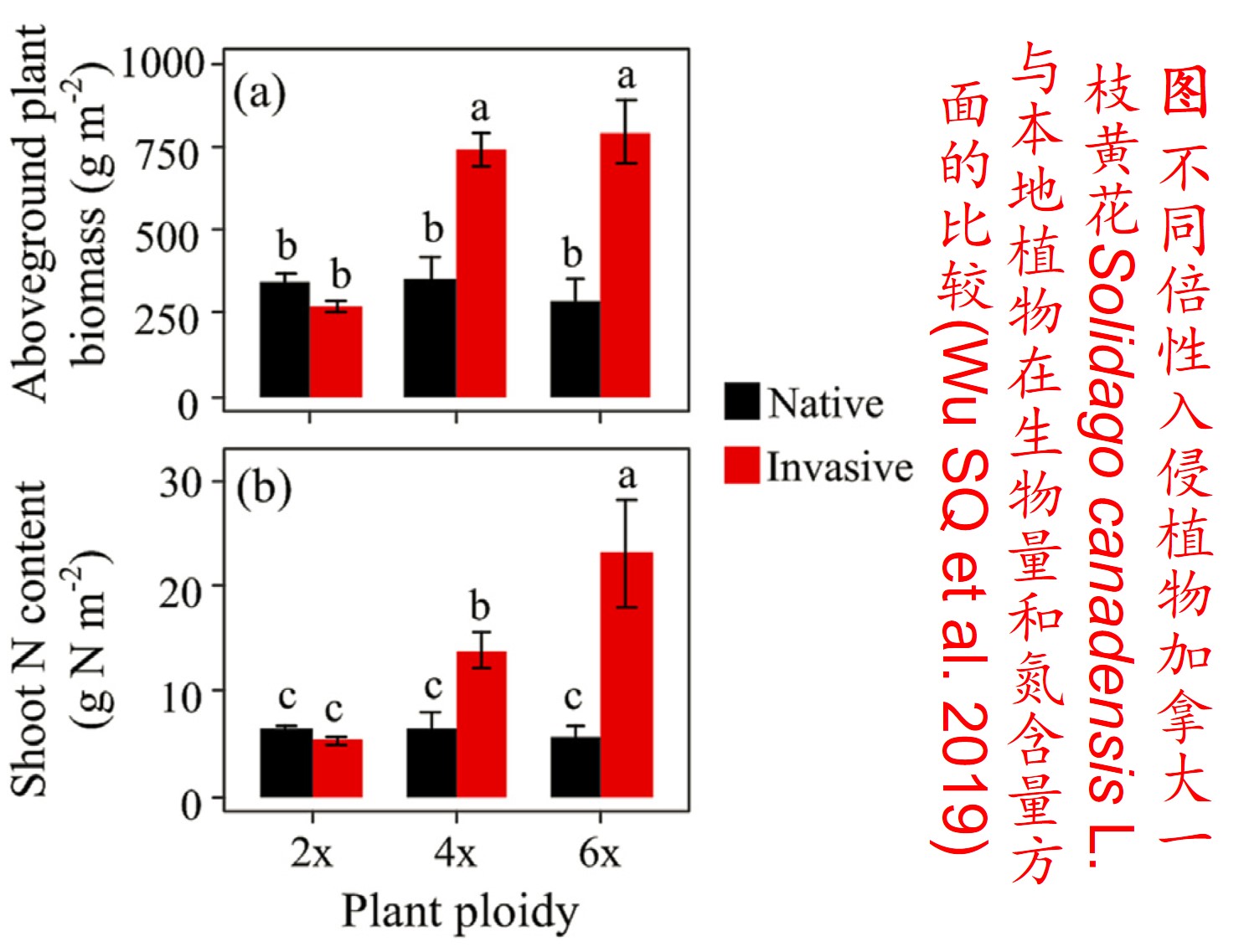

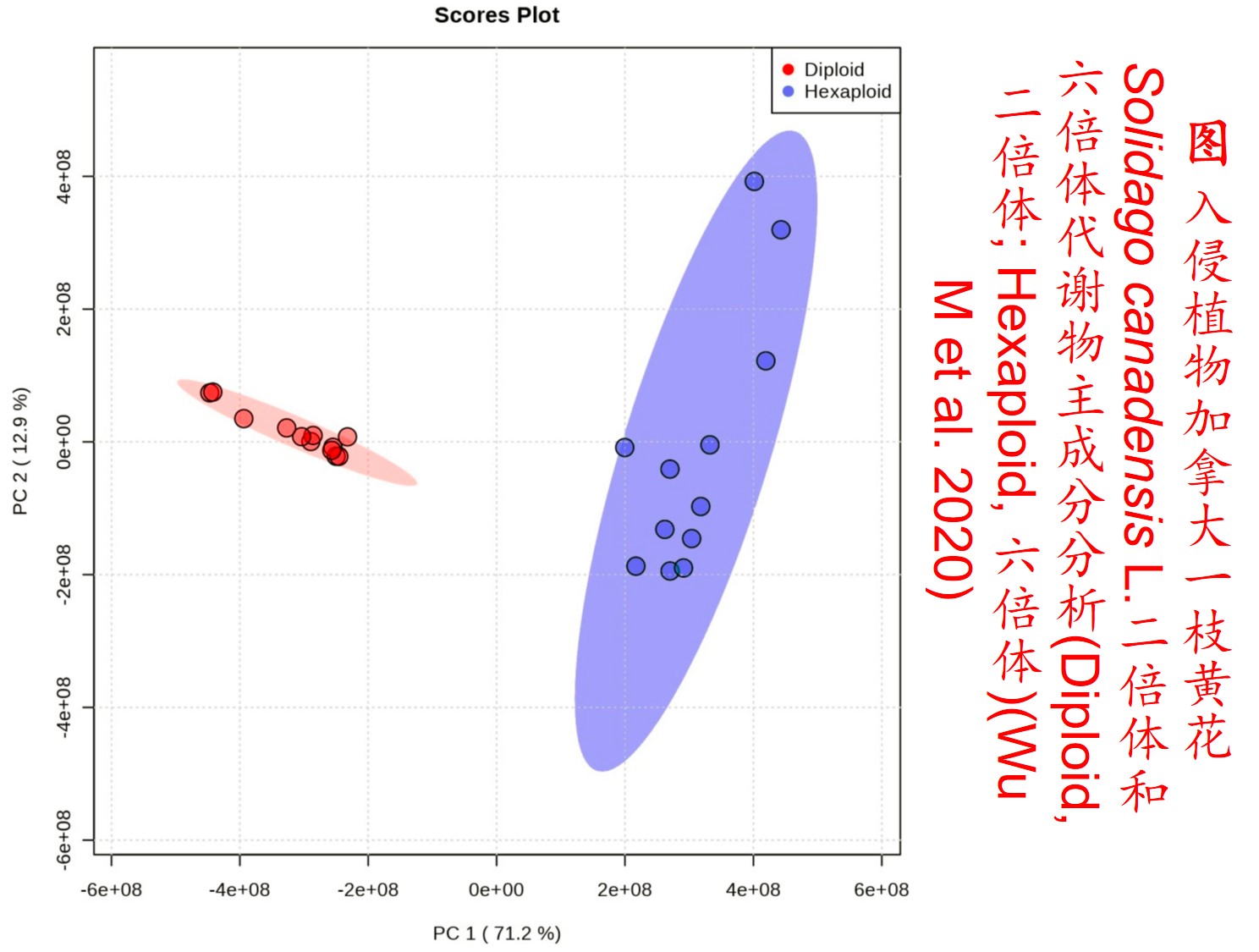
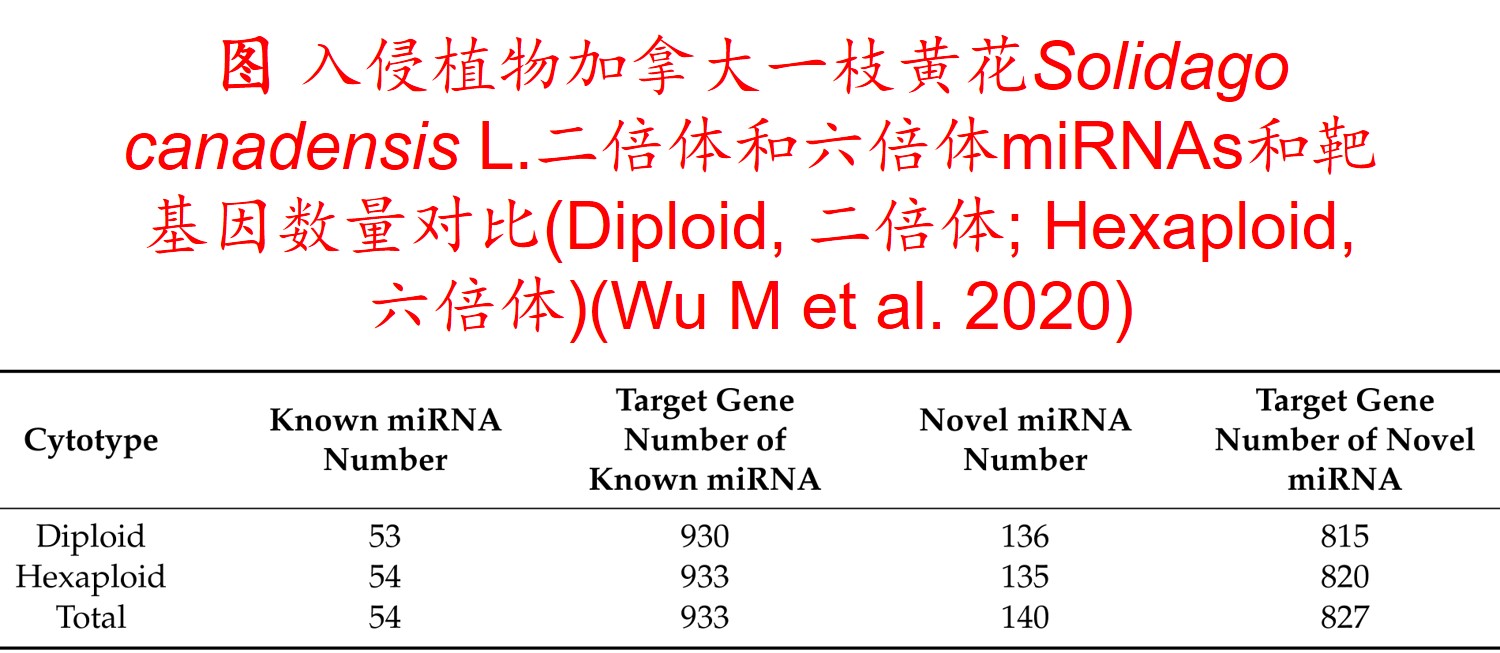

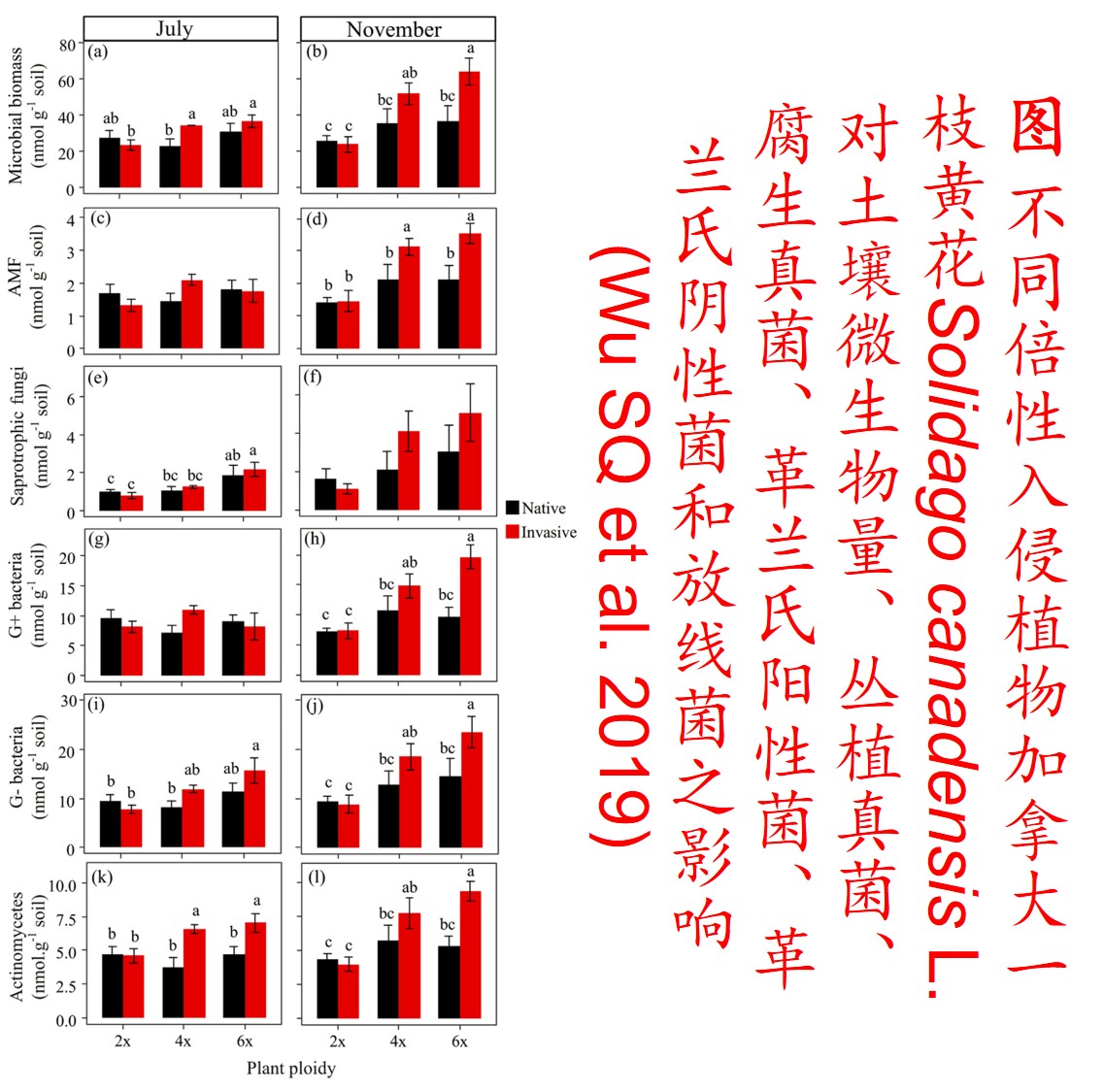
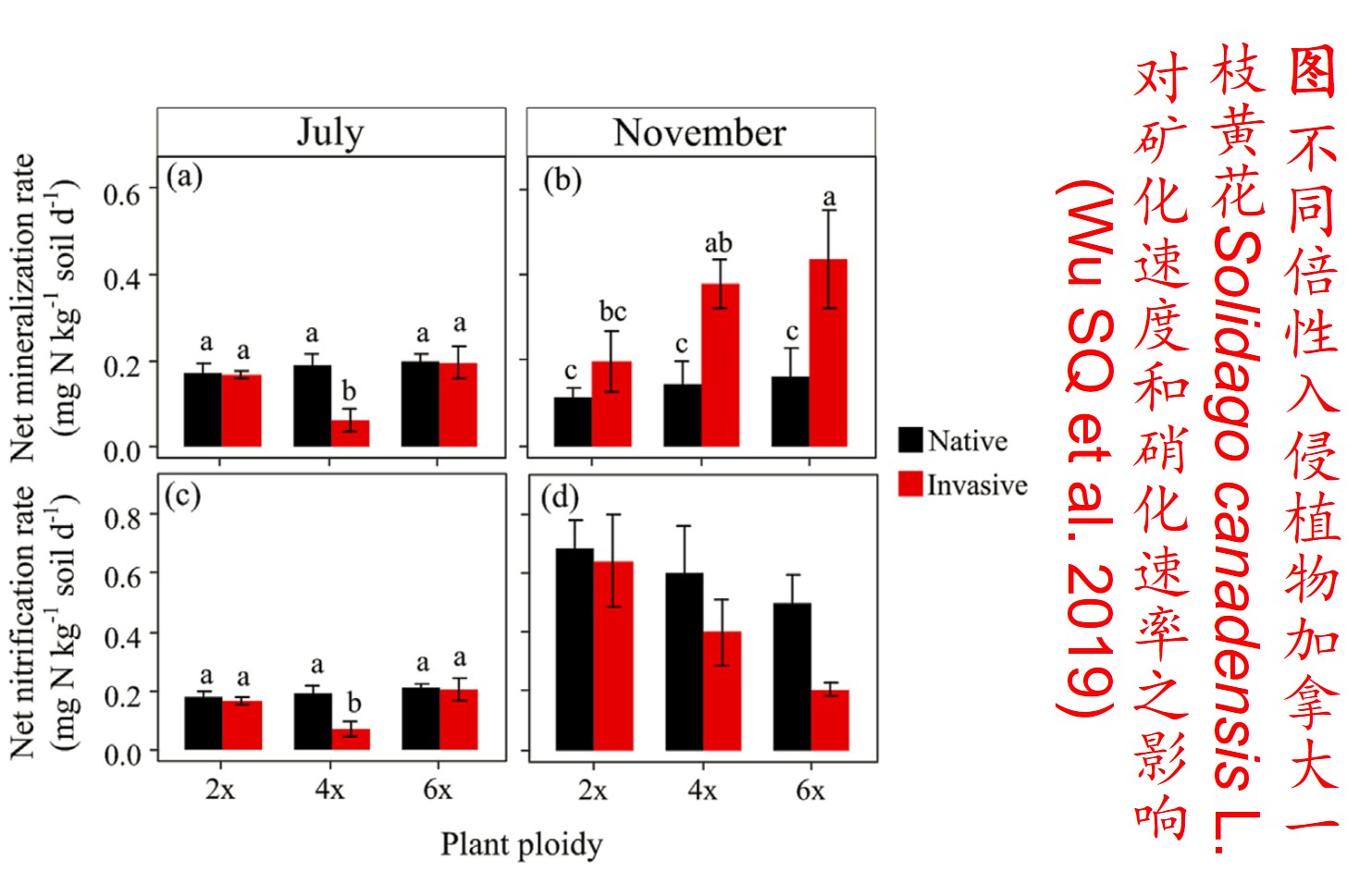
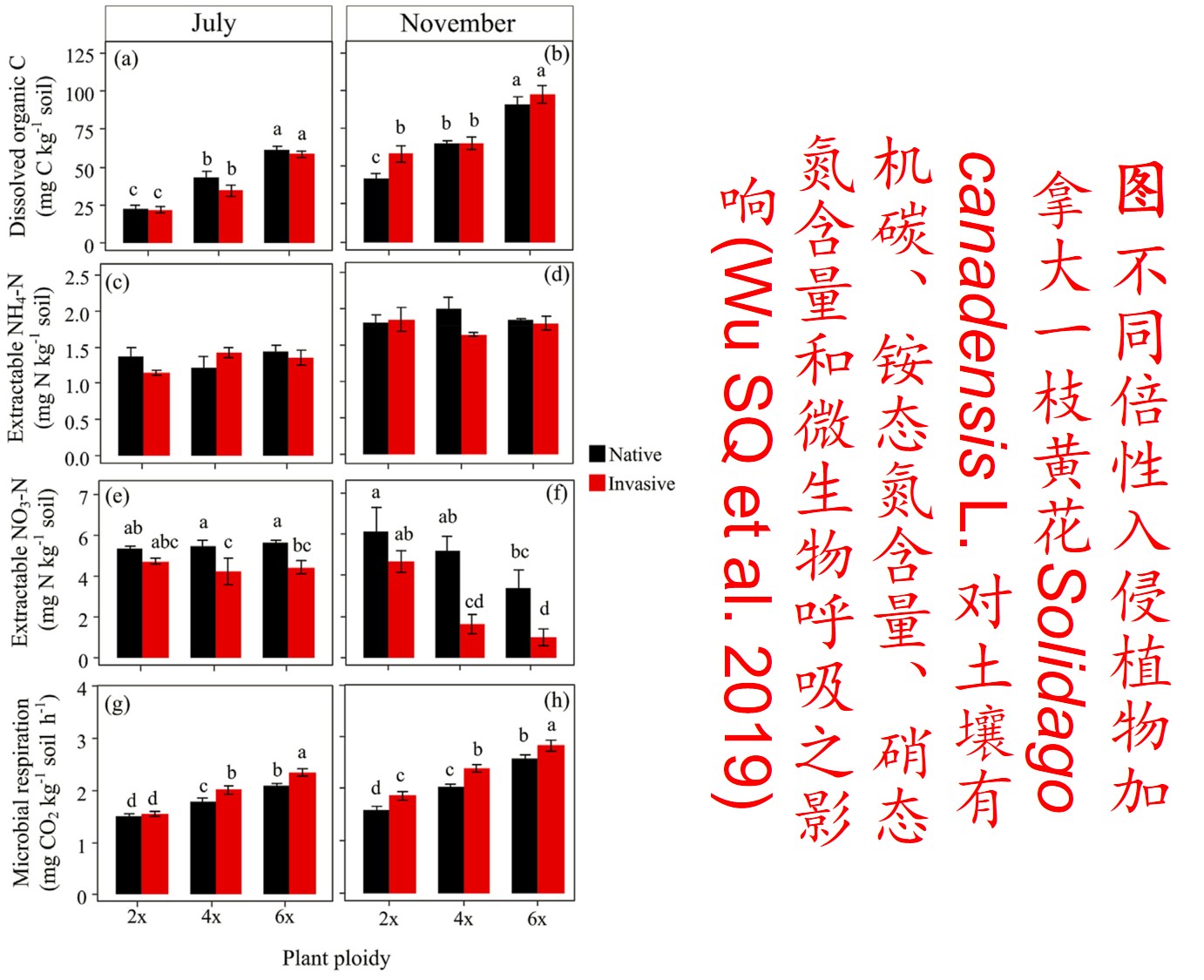
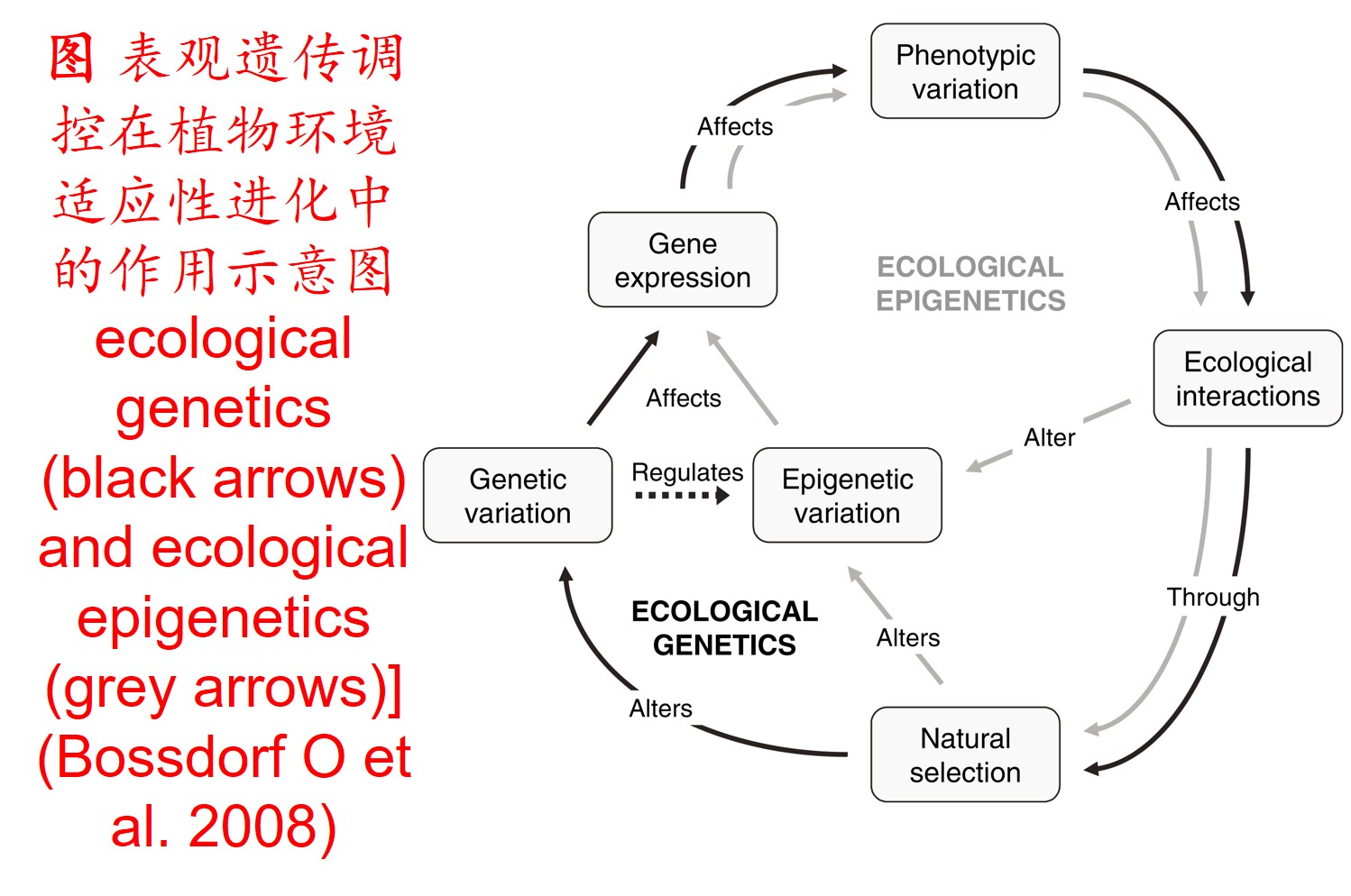
5. 表观遗传调控(Epigenetic regulation):
表观遗传(Epigenetics)在调控和驱动生物在新生境适应性进化过程中扮演着重要角色(生物的表观遗传可参阅http://blog.sciencenet.cn/blog-565899-1221373.html)。
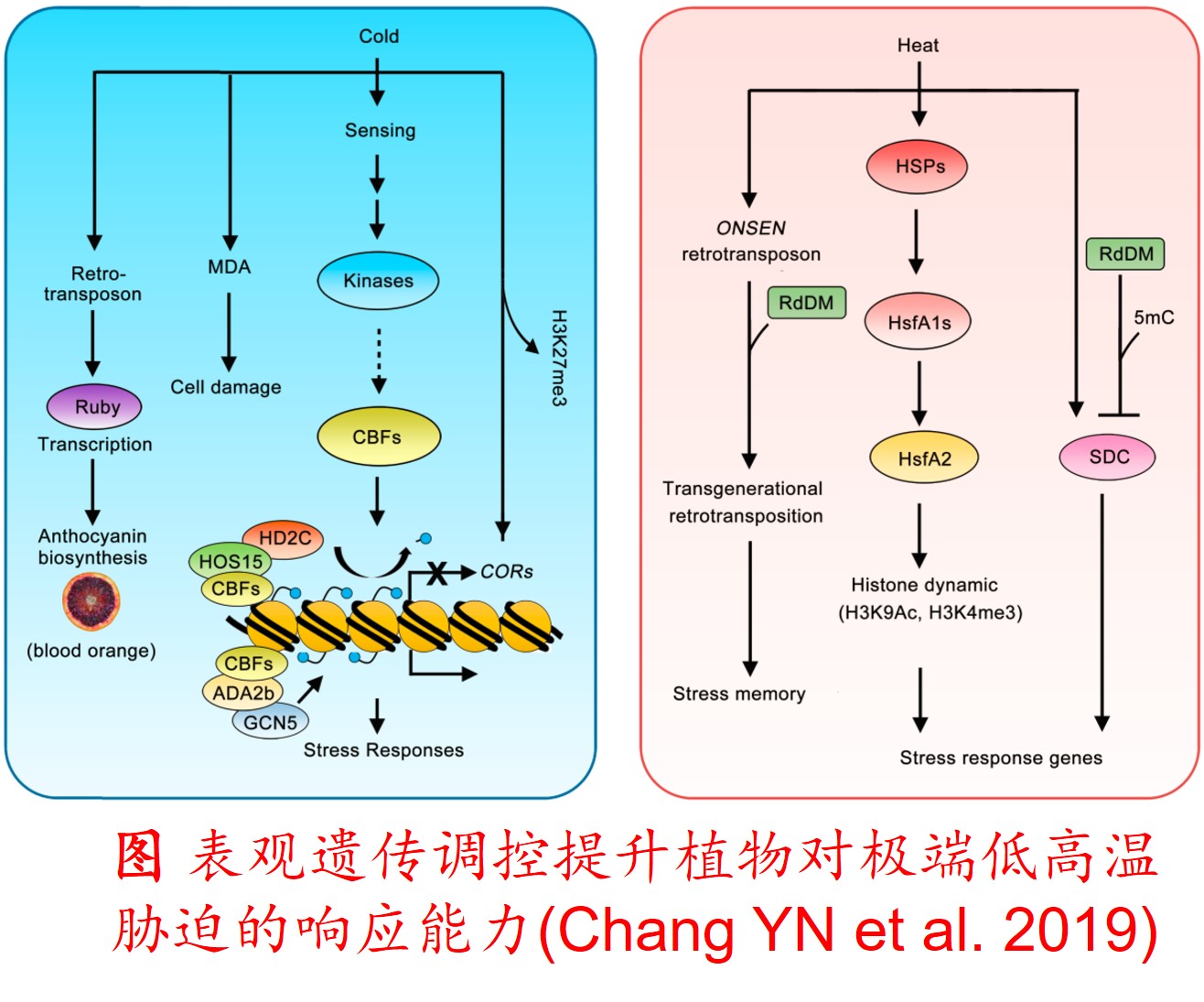
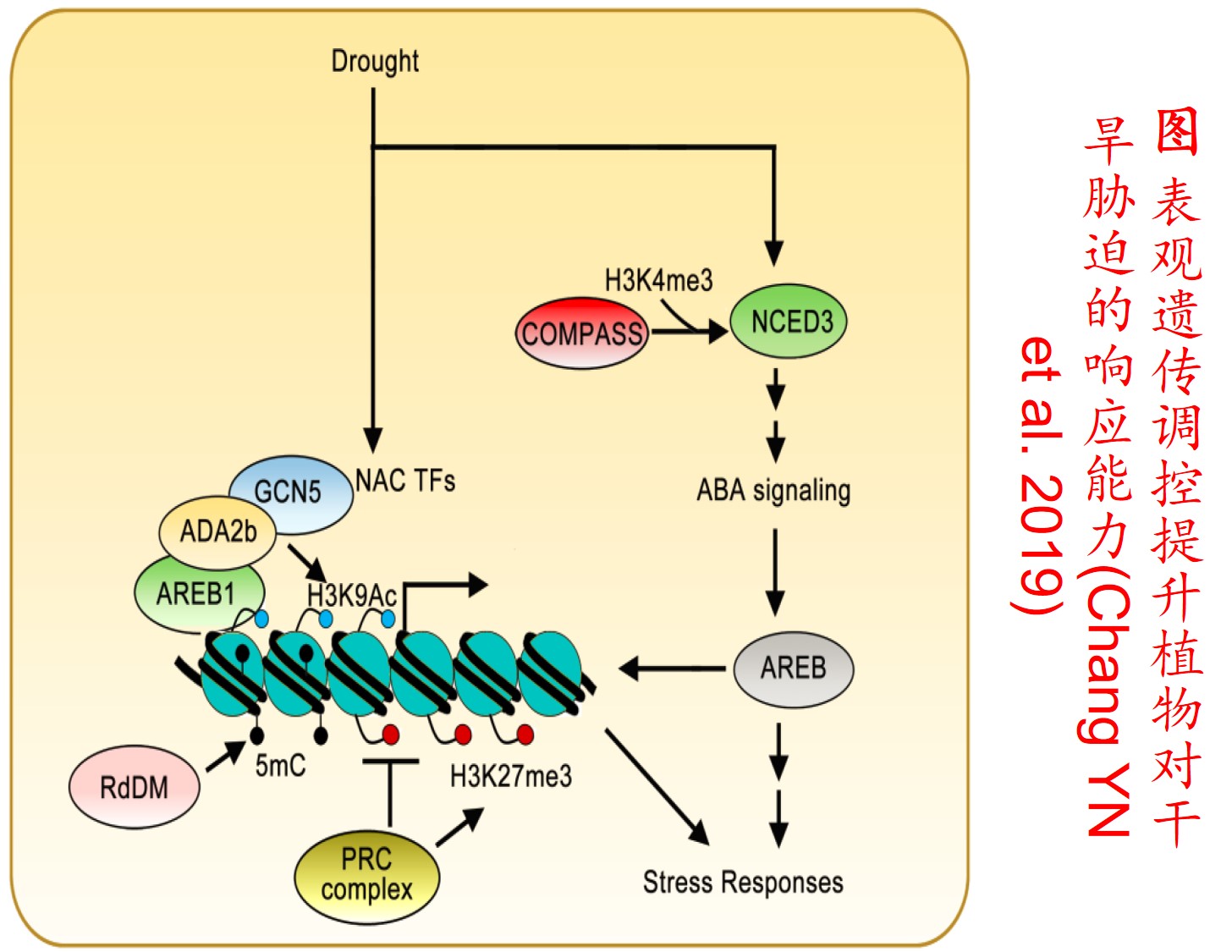
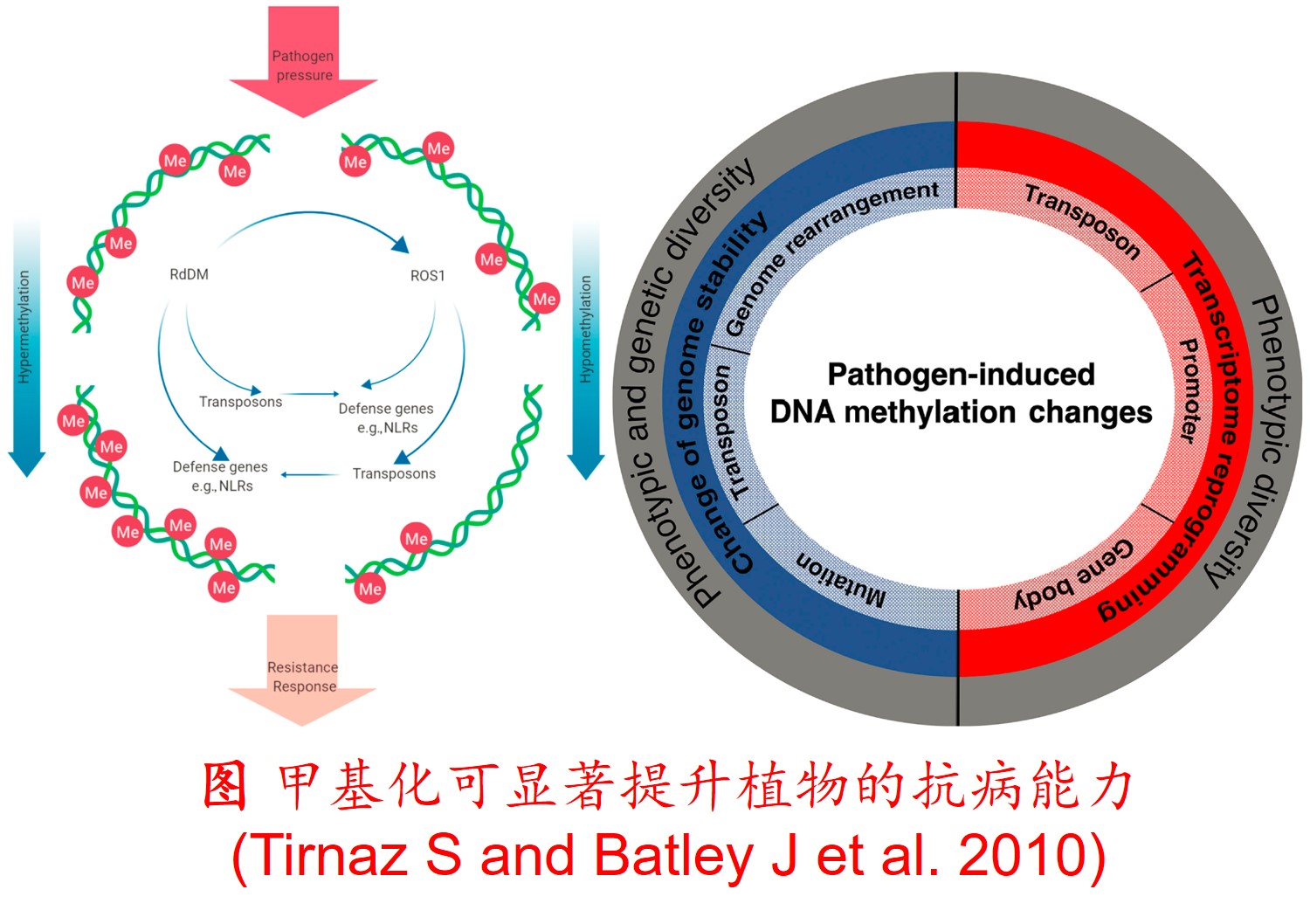
在分子遗传机制方面,表观遗传调控主要是通过DNA甲基化、组蛋白修饰和非编码RNA调控等途径来完成,而DNA甲基化是表观遗传调控最为关键的组成部分之一,且DNA甲基化(水平通常为6%至30%)普遍存在于高等植物中(主要发生在CpG和CpNpG位点上),一般情况下主要形式为DNA序列5’端胞嘧啶甲基化,以用于维持基因组的稳定性和调节基因表达。
此外,植物可通过甲基化、去甲基化和从头甲基化作用等诸多形式在不改变基因组DNA系列的前提下调控其基因表达,进而影响其生理生态特性及其对环境的适应性进化,尤其是植物在胁迫环境下可通过调节DNA甲基化水平的路径调控其基因表达,从而快速适应不利的环境。
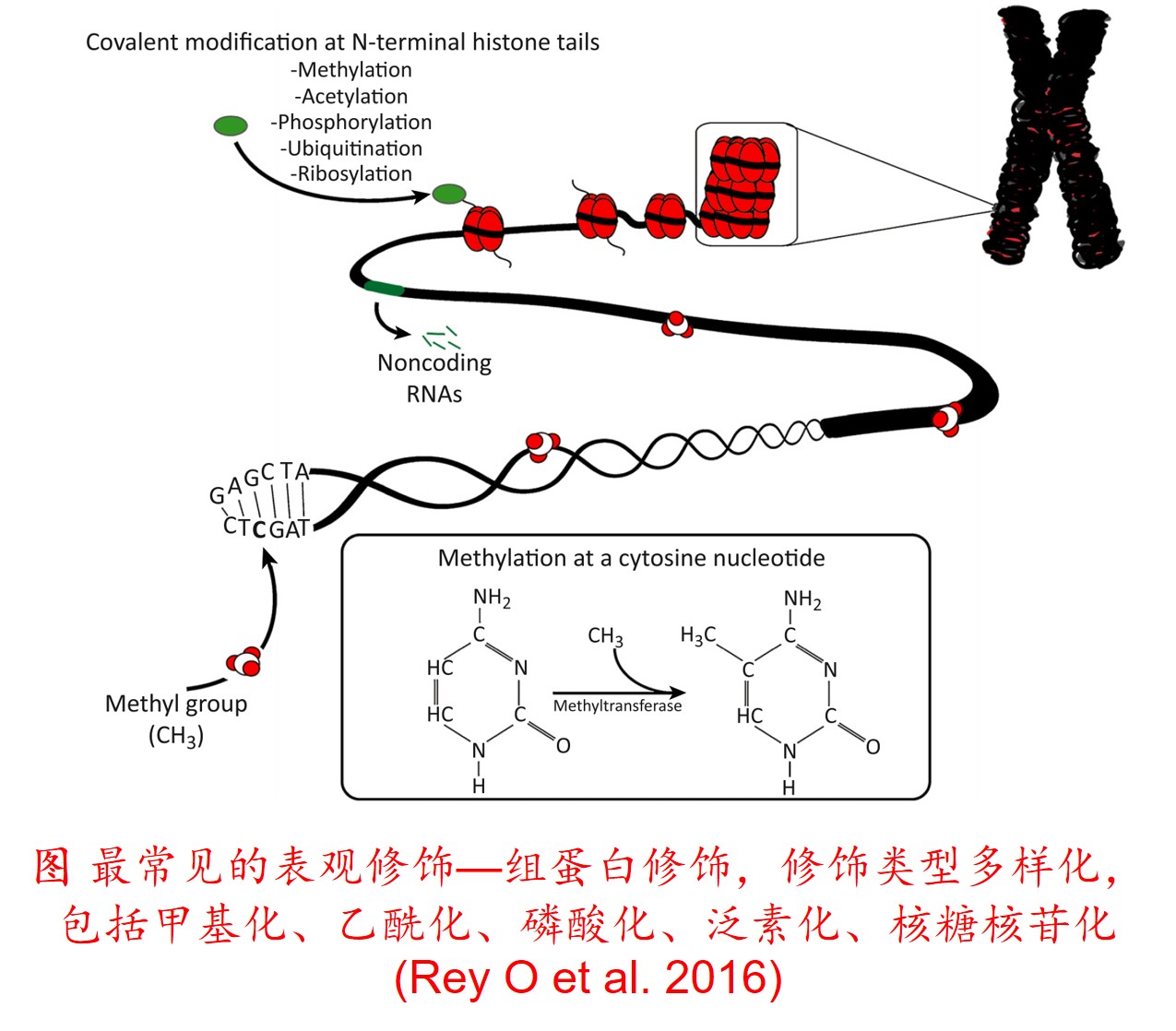
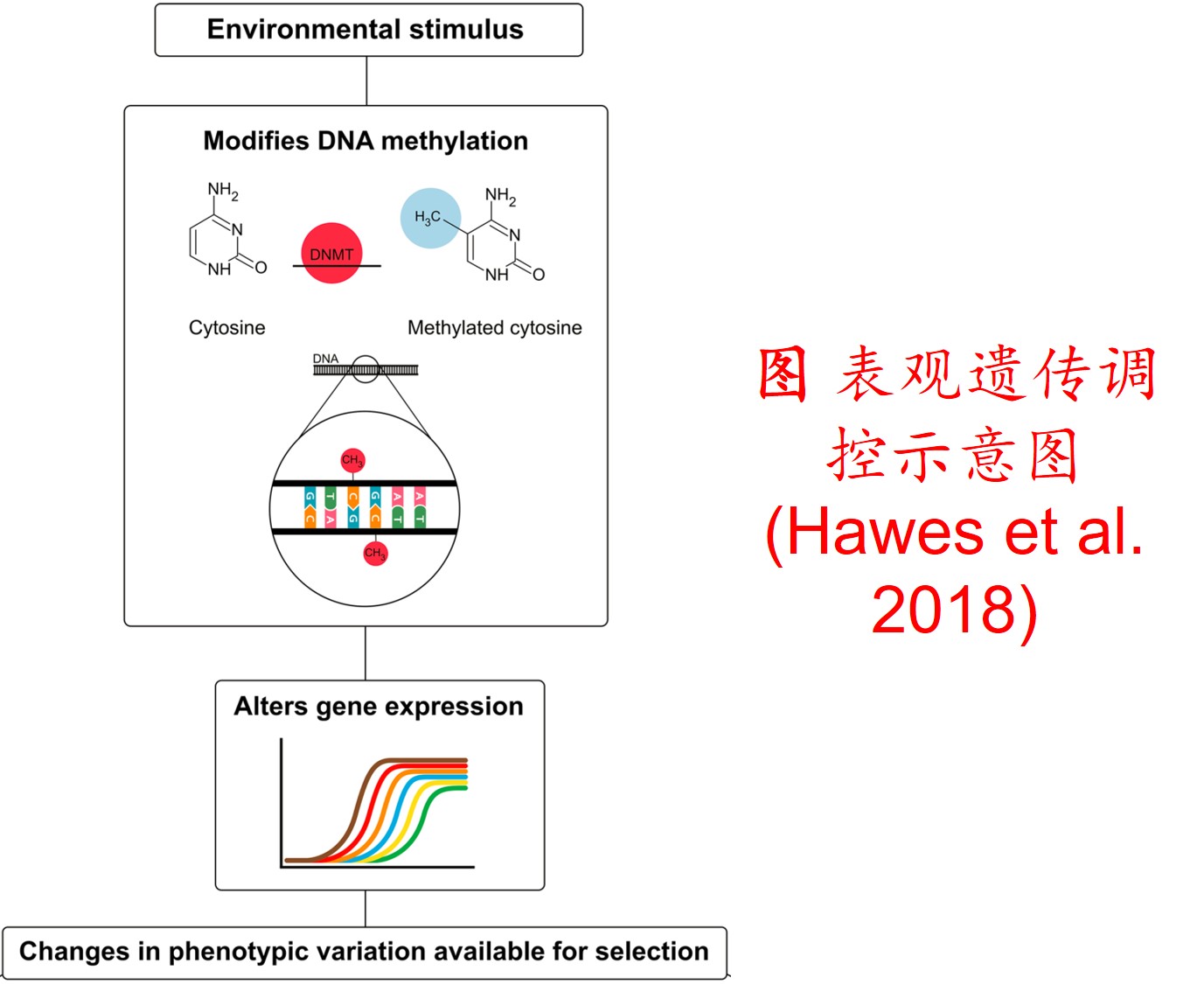


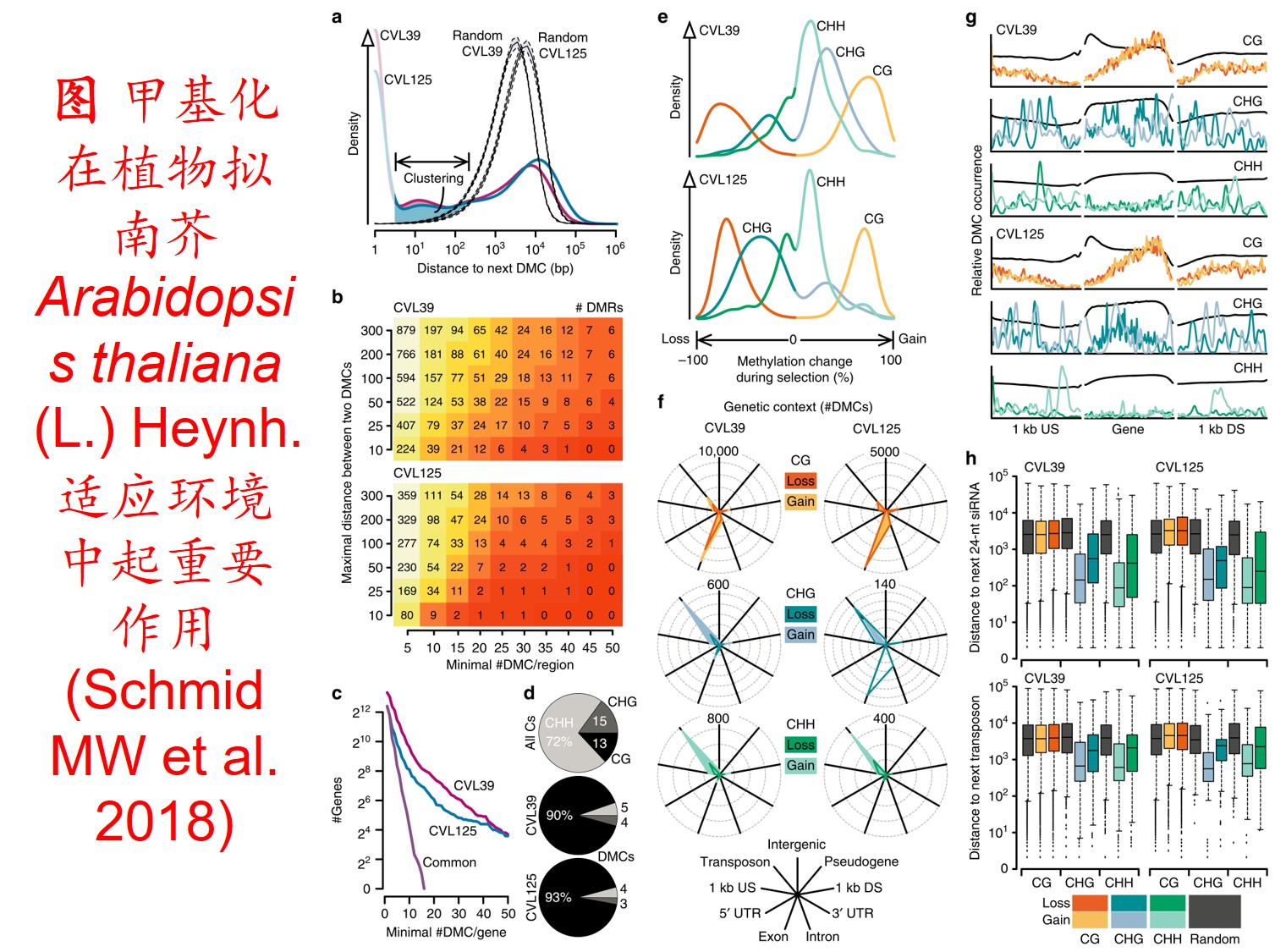
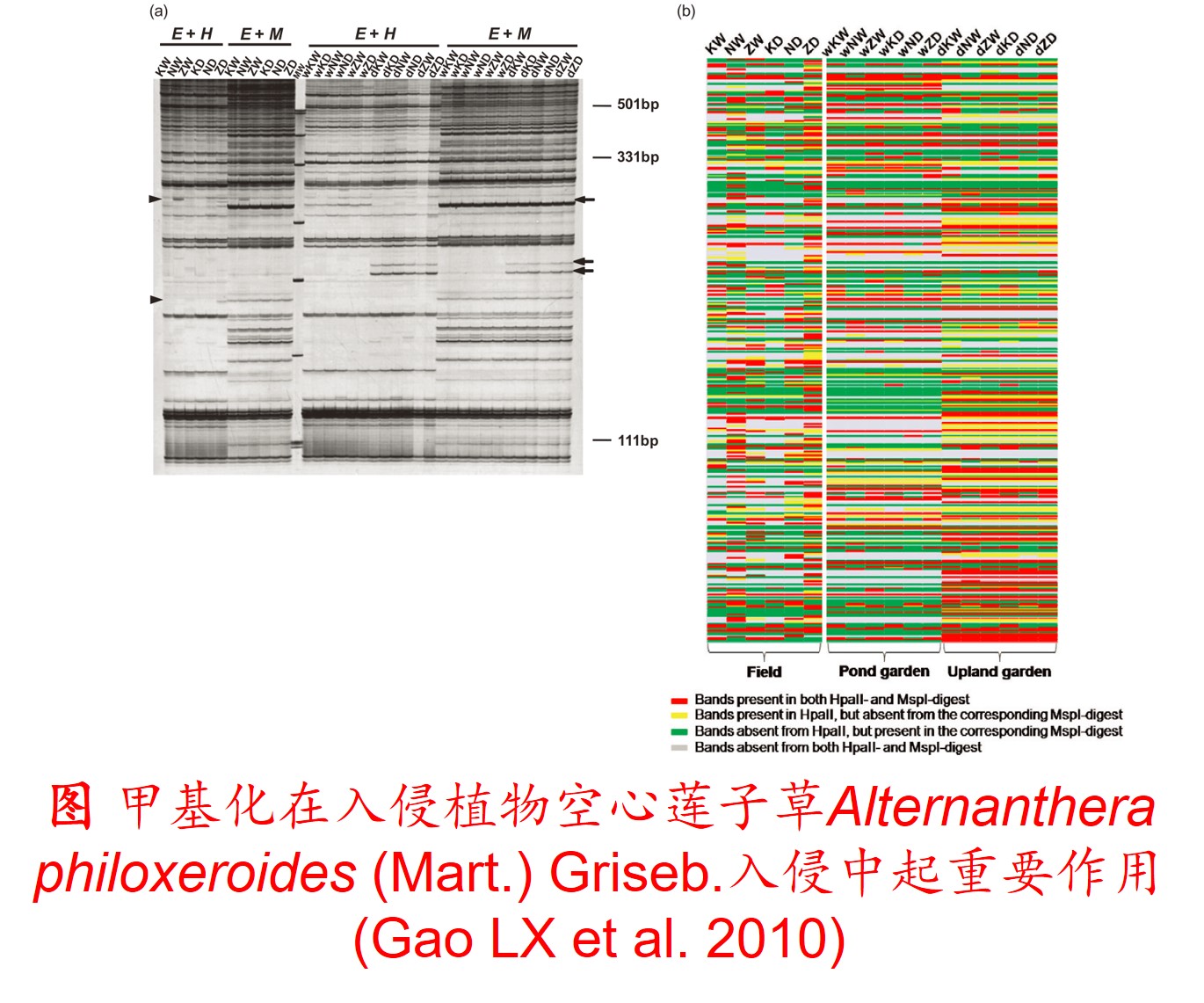
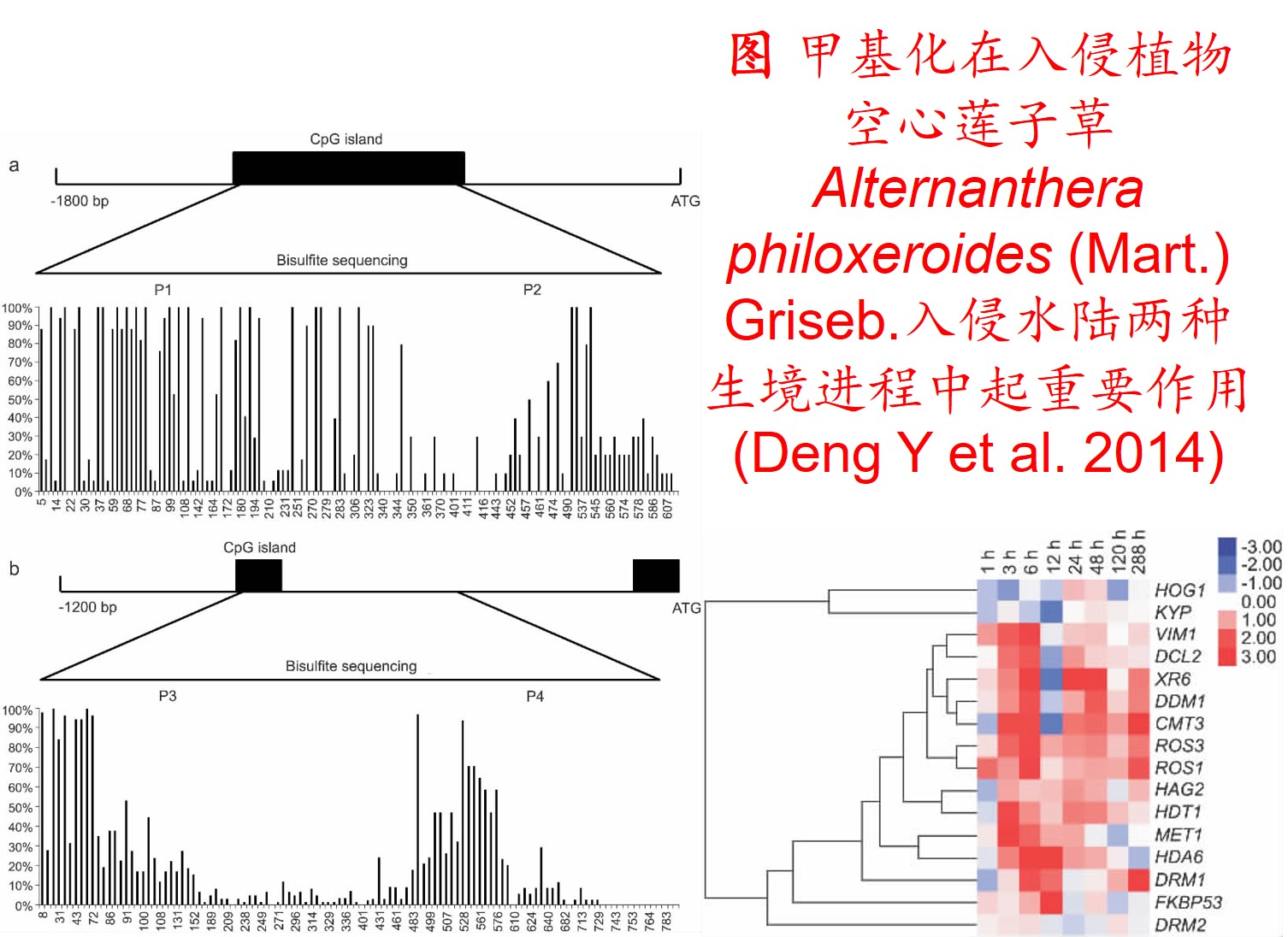
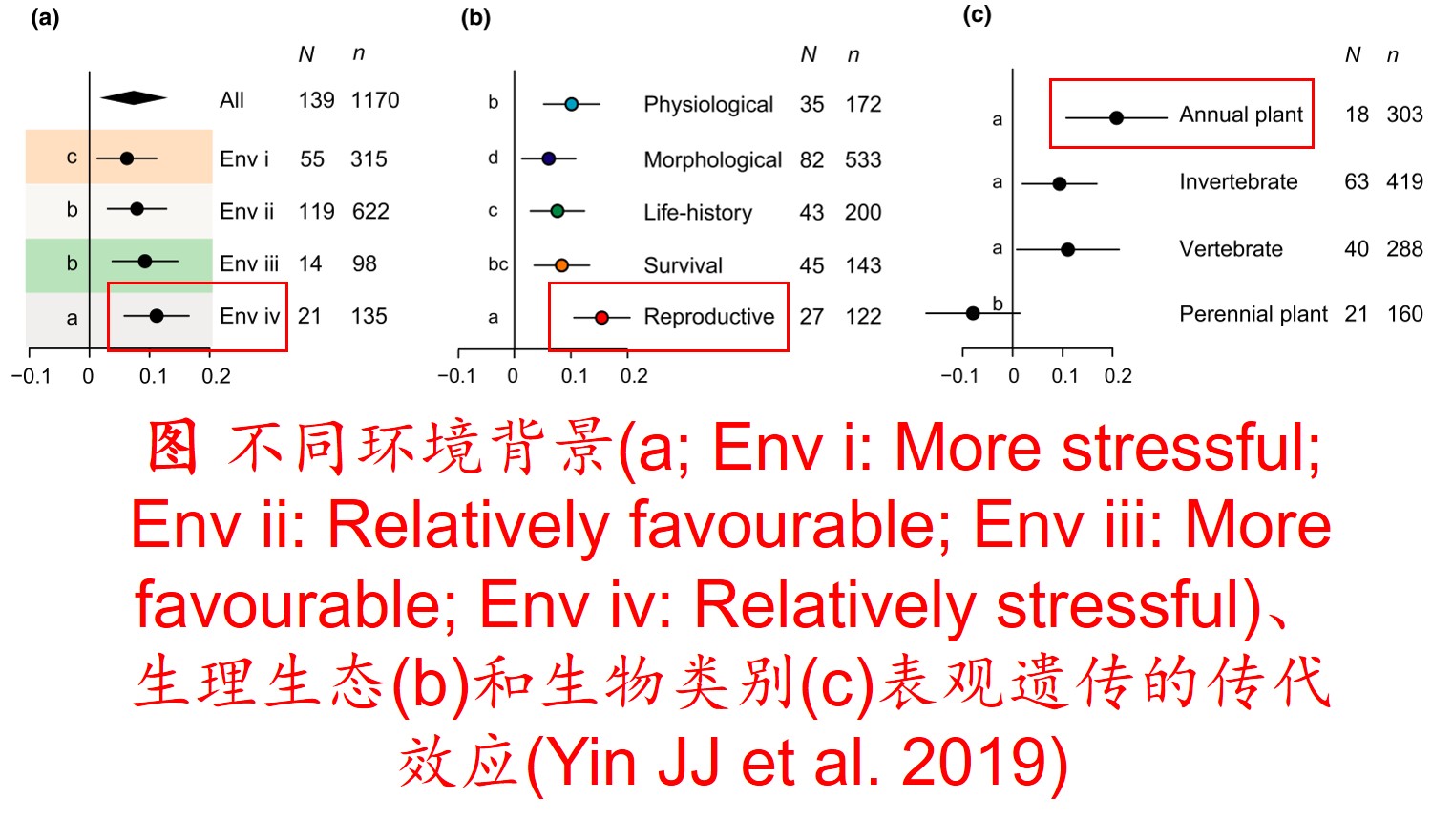

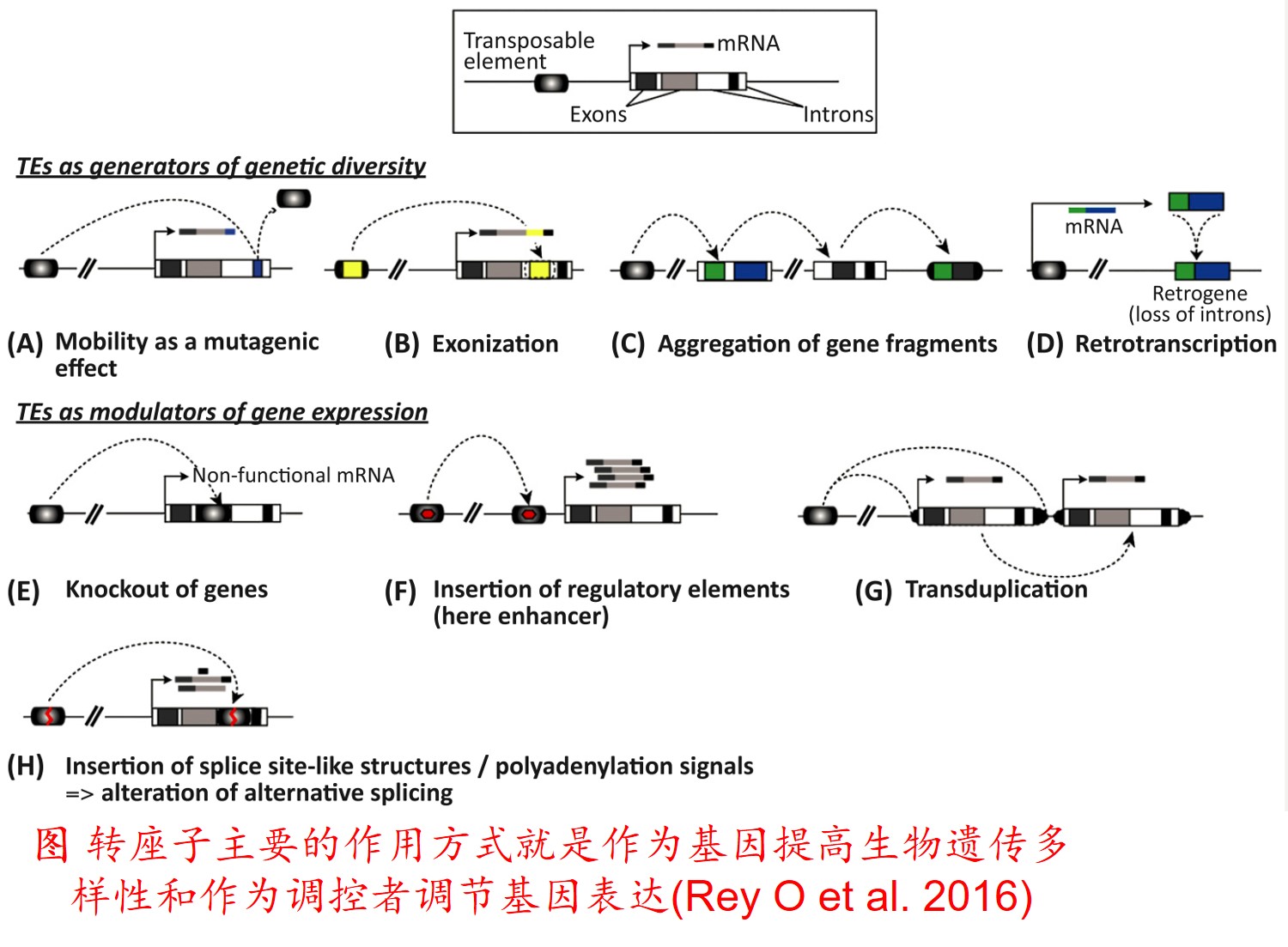
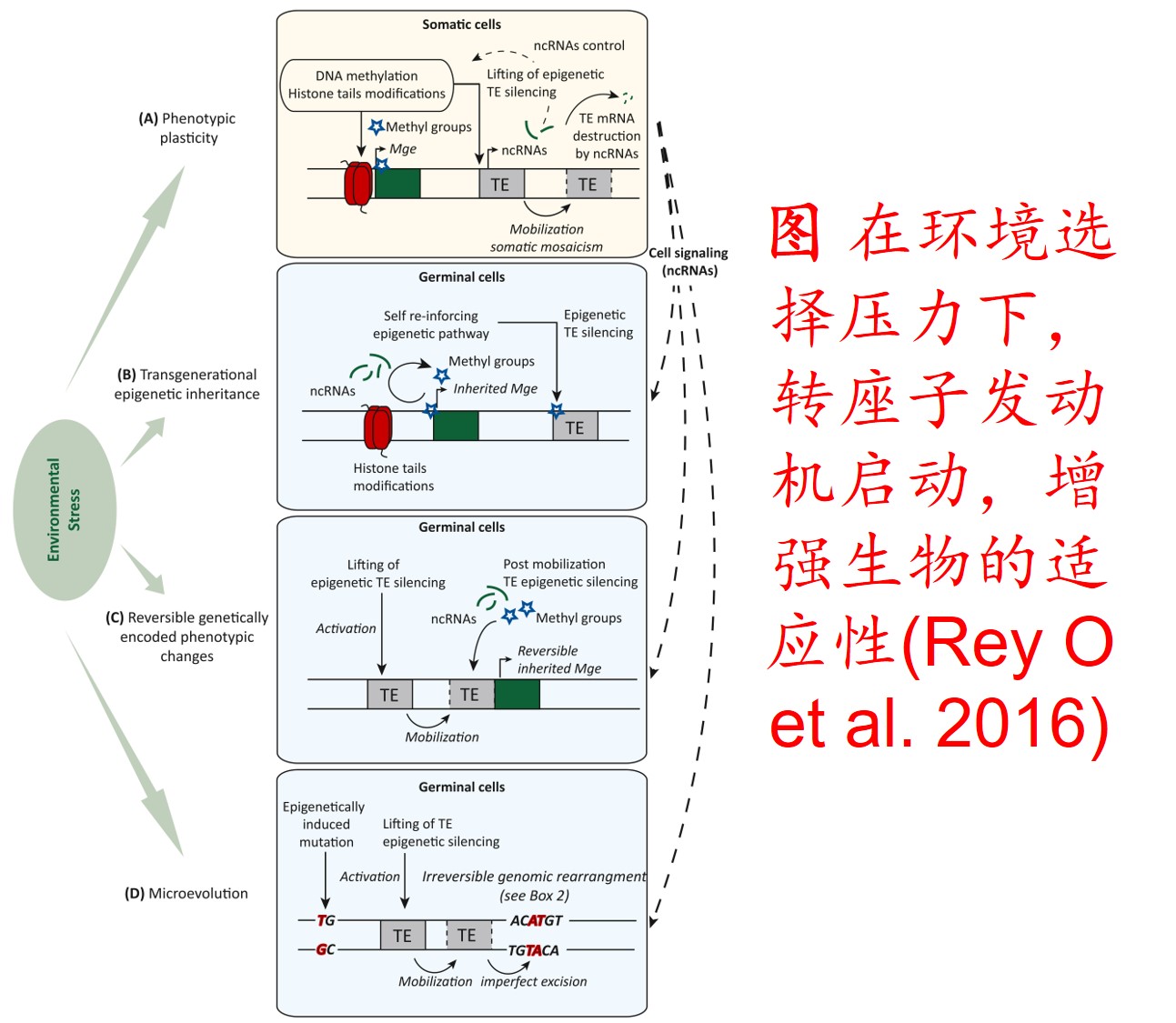
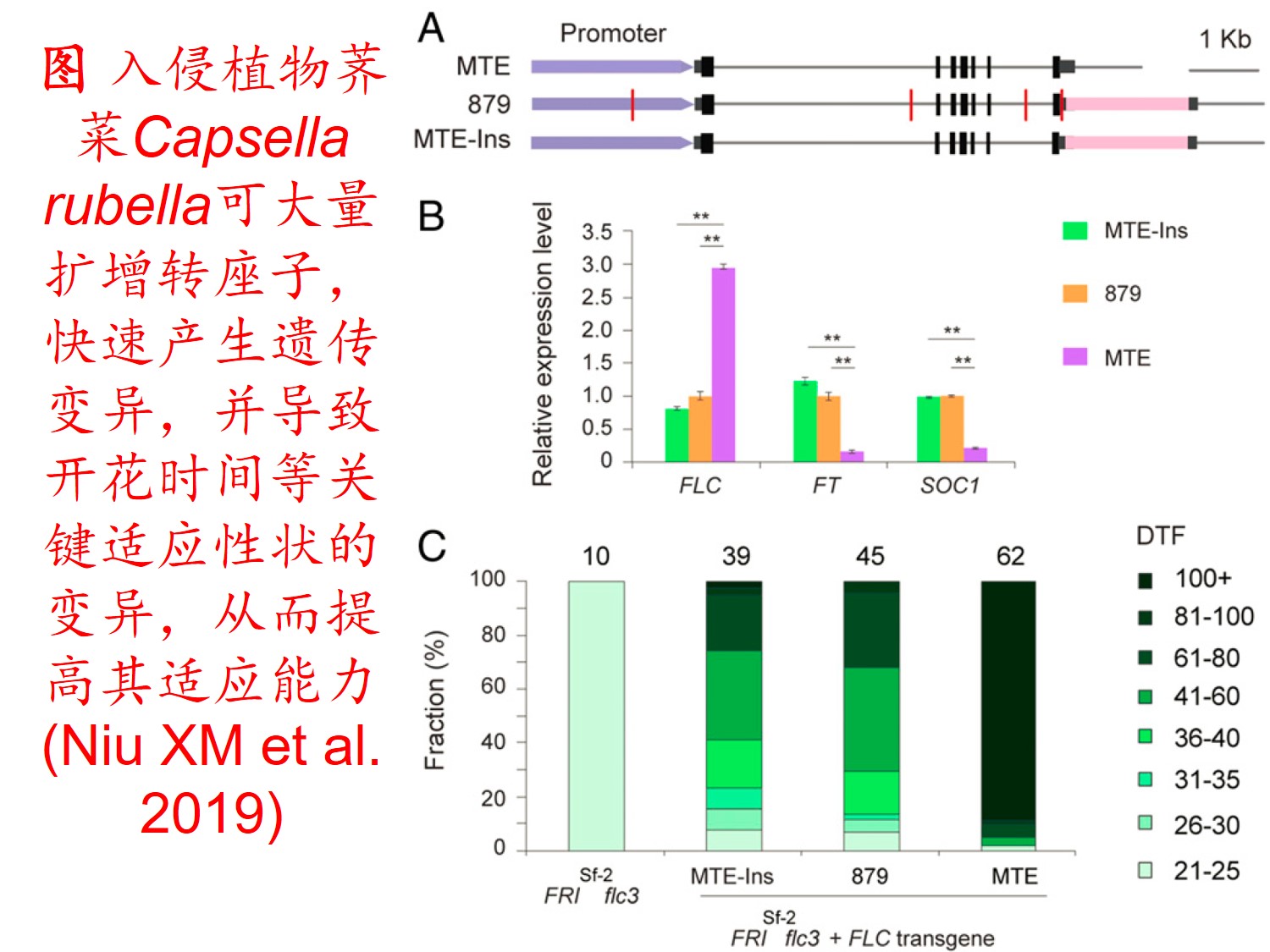
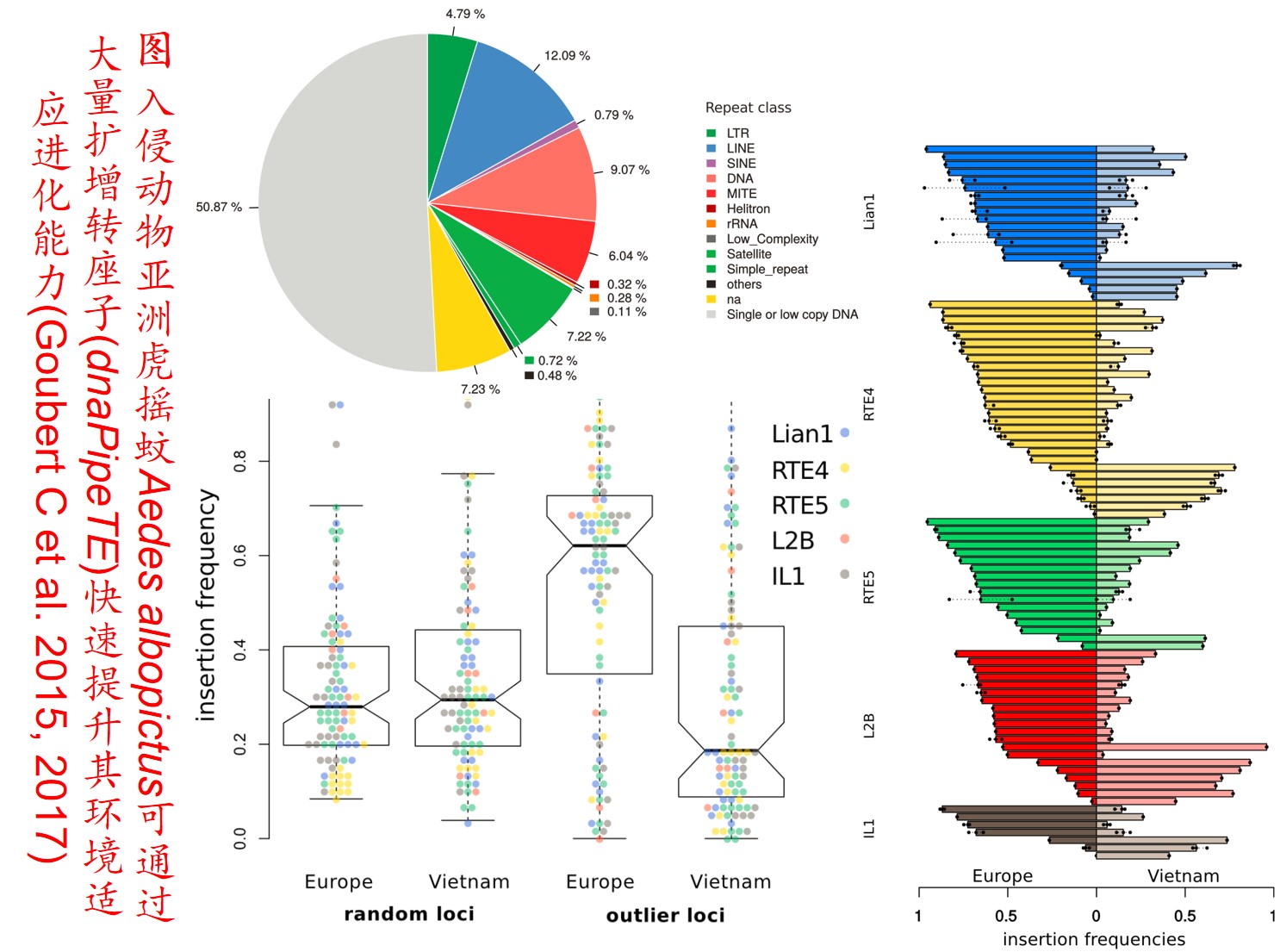
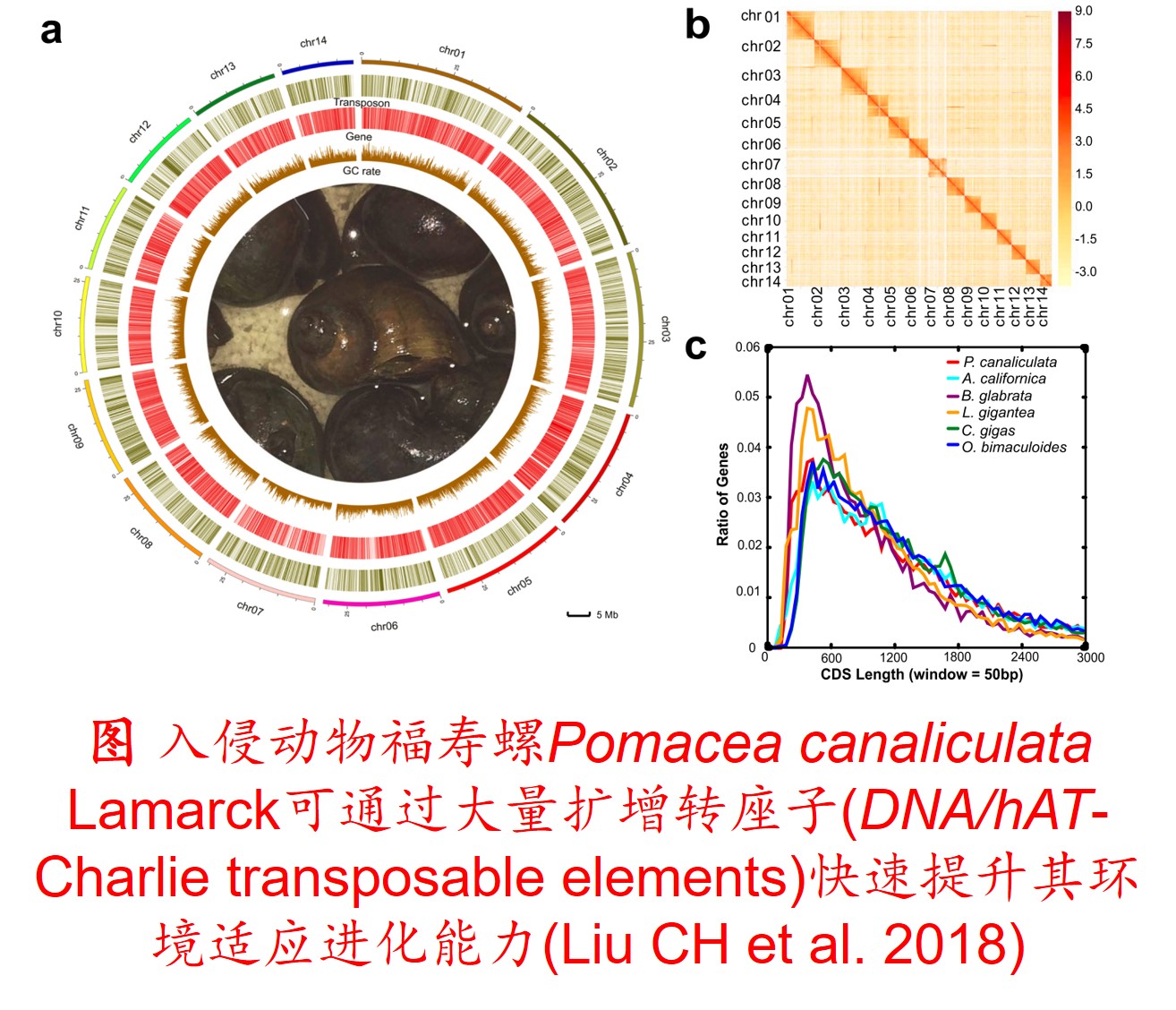
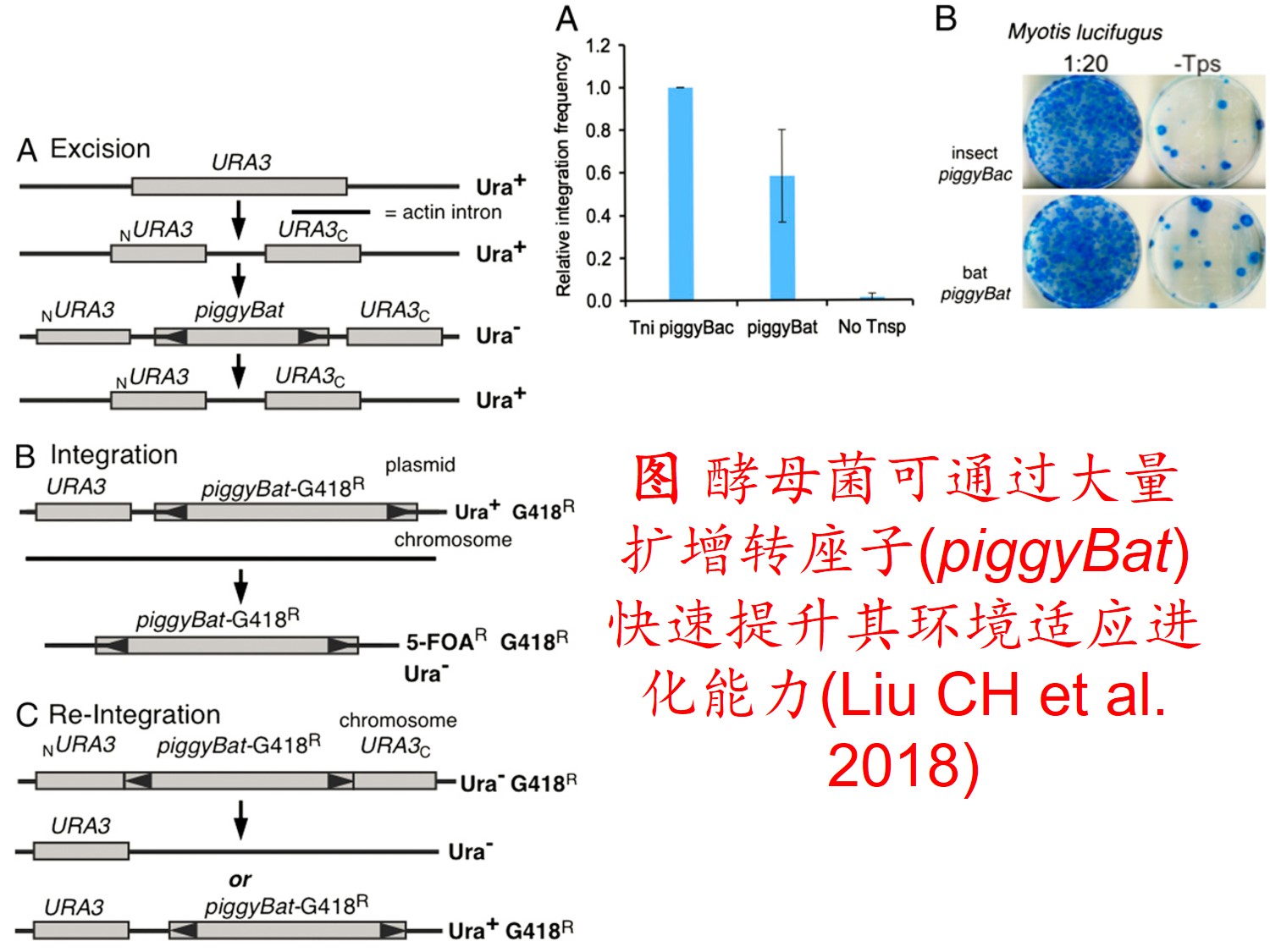
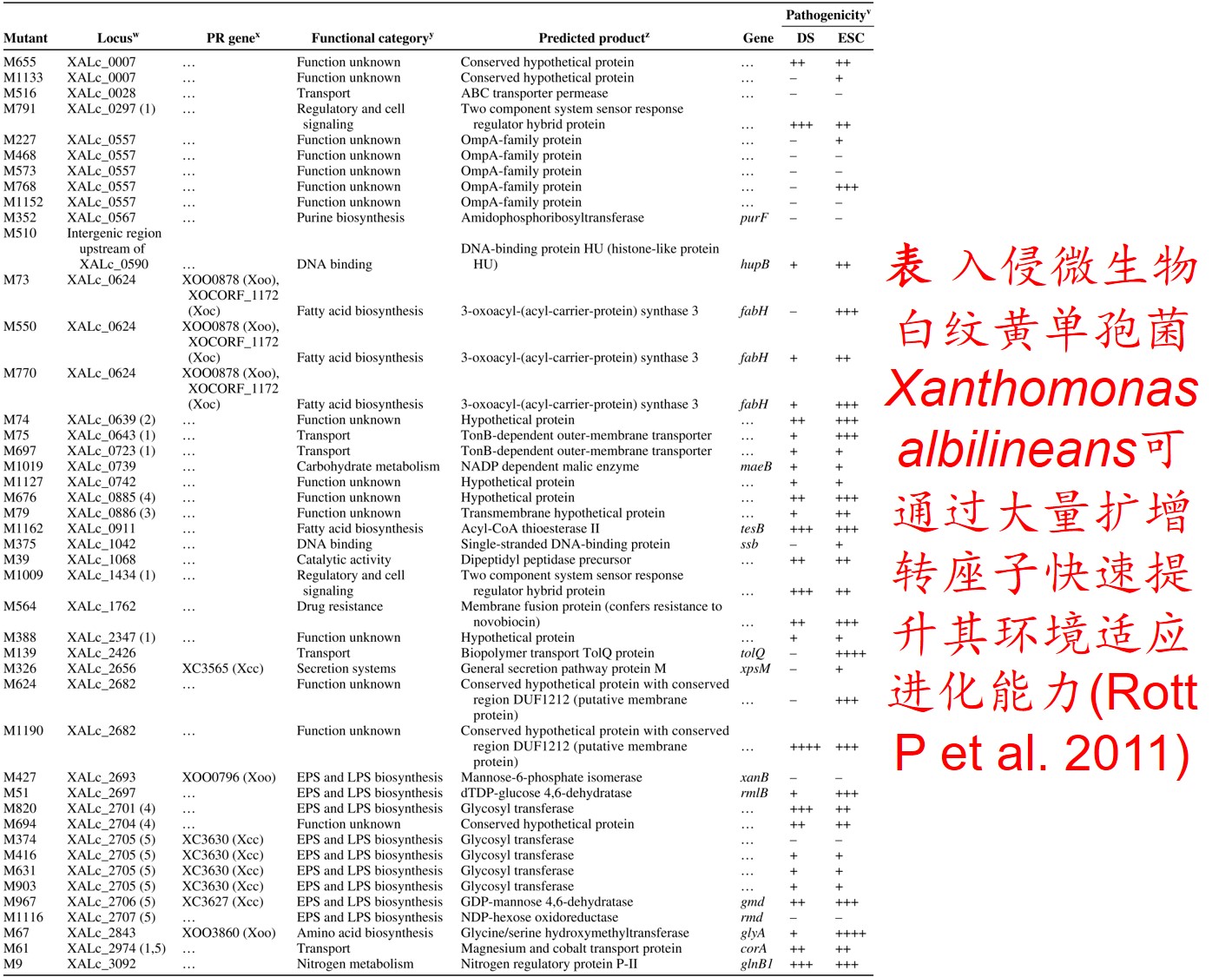
6. 转座子(Transposon)大量扩增:
转座因子或转座子是一类在很多生物体内发现的可移动的遗传因子。 一段基因可从原位上单独复制或断裂下来,环化后插入另一位点,并对其后的基因起调控作用,此过程称转座。这段序列称跳跃基因或转座子,可分插入序列(Is因子)、转座(Tn)和转座phage。
而遗传多样性很低的物种可通过转座子大量扩增,快速产生遗传变异,并导致关键适应性状的变异,从而提高其适应新生境的能力。
注:囿于个人水平之限,错误在所难免。敬请批评指正!此外,本博文图片和部分文字介绍均源于网络,版权归原作者和(或)出版商,仅用于学术传播等非商业行为。特在此致谢!
生物入侵机制http://blog.sciencenet.cn/blog-565899-1206570.html
入侵植物的入侵竞争力和生境的可入侵性量化方法http://blog.sciencenet.cn/blog-565899-1219837.html
群落多样性与其可入侵性的关系的悖论http://blog.sciencenet.cn/home.php?mod=space&uid=565899&do=blog&id=1211044
植物群落物种共存机制悖论http://blog.sciencenet.cn/home.php?mod=space&uid=565899&do=blog&id=1208621
入侵植物与氮互作存在问题及未来研究展望http://blog.sciencenet.cn/home.php?mod=space&uid=565899&do=blog&quickforward=1&id=1209142
生物入侵途径http://blog.sciencenet.cn/blog-565899-1203613.html
生物入侵现状http://blog.sciencenet.cn/blog-565899-1204926.html
生物入侵进程http://blog.sciencenet.cn/home.php?mod=space&uid=565899&do=blog&quickforward=1&id=1207475
中国入侵植物名单和名录http://blog.sciencenet.cn/blog-565899-1196601.html
中国入侵植物名录和入侵克隆植物名录http://blog.sciencenet.cn/blog-565899-961990.html
中国水生入侵植物名录http://blog.sciencenet.cn/blog-565899-973994.html
中国外来入侵植物图谱http://blog.sciencenet.cn/blog-565899-1185806.html
中国&江苏外来入侵植物名录http://blog.sciencenet.cn/home.php?mod=space&uid=565899&do=blog&quickforward=1&id=1212281
惨不忍睹的生物入侵http://blog.sciencenet.cn/blog-565899-1145336.html
世界上入侵力最强的100种外来入侵植物http://blog.sciencenet.cn/home.php?mod=space&uid=565899&do=blog&id=1205853
BBCH-scale (Weed)http://blog.sciencenet.cn/home.php?mod=space&uid=565899&do=blog&id=1205474
https://wap.sciencenet.cn/blog-565899-1225699.html
上一篇:一枝江南春意浓
下一篇:陌上花开-木瓜