博文
活性氧:棘手的抗肿瘤和促肿瘤双面分子
 精选
精选
||
活性氧具有双面性,一方面具有信号作用,一方面具有毒性。一般学者和公众对活性氧的毒性往往比较认可甚至夸大,但往往忽视活性氧的信号或功。生物体系中,几乎没有绝对的有毒和有用物质,有毒物质往往会有正面作用,毒性越大的东西越能发挥更大作用,有用的东西如果浓度太高往往有害,有毒和有用是相对的,一般决定于剂量。例如葡萄糖是维持机体正常功能的重要因素,不足可以发生低血糖,危机生命,而血糖水平高就是糖尿病。再如二氧化碳有毒,但是没有二氧化碳就无法有效刺激呼吸,急性高原病虽然起因是缺氧,但重要因素是体内二氧化碳不足造成。氧气是必须的,但高浓度氧气对生物是有很大毒性的,过高和不足都是不好的,只有适当的氧浓度才适合生命活动。活性氧和氧气、葡萄糖、二氧化碳这些生物分子都具有类似特征。
以下内容主要选择于2014年Current Biology 24, R453–R462,的一篇综述ROS Function in Redox Signaling and Oxidative Stress。
氧气是需氧生物不可缺少的物质,在体内获得4个电子可以变成水,但如果只获得一个电子,则变成不完全还原的超氧阴离子,细胞内形成超氧阴离子的主要有线粒体途径和NADPH氧化酶模式。超氧阴离子属于离子,不容易在细胞内扩散,生物在进化过程中获得催化超氧阴离子变成过氧化氢的SOD,因此在细胞内过氧化氢的浓度是超氧阴离子的1000倍,一般来说,如果不特别声明,细胞内活性氧就是指过氧化氢。过氧化氢在体内产生主要依靠SOD催化超氧阴离子,而降解过氧化氢的酶种类比较多,主要包括过氧化氢酶、谷胱甘肽过氧化物酶和过氧化物酶三种催化方式。在疾病过程中,一些结合在蛋白中的金属离子如铁释放为游离状态,可以和过氧化氢发生feton反应,产生羟基自由基,羟基自由基是自然界中氧化作用最强的物质之一,在生物体系内半衰期为1纳秒,一旦产生,就会和蛋白质、核酸和脂类物质发生氧化反应,造成氧化损伤。由于羟基自由基的毒性极强,而且没有特异性清除方法,细胞只有通过多种措施限制金属离子释放。在生物体系内,虽然活性氧的种类很多,超氧阴离子、过氧化氢和羟基自由基最为典型,分别代表活性氧的生成,信号效应分子和毒性分子。其他如一氧化氮、亚硝酸阴离子、氧化型抗坏血酸和各种脂质过氧化物等都属于活性氧,但都没有这三种典型,因此在文献中,学者研究的对象主要是针对超氧阴离子、过氧化氢和羟基自由基。
活性氧的信号比较典型的模式是过氧化氢对半胱氨酸的氧化调节。半胱氨酸含有一个巯基Cys–SH,在生理条件下蛋白质中的半胱氨酸巯基会水解出一个氢离子,成为巯基阴离子Cys–S-,巯基阴离子对氧化比巯基更敏感。当细胞内过氧化氢水平增加时,能将巯基阴离子氧化为半胱次磺酸(Cys–SOH),这一变化能引起蛋白发生变构,影响蛋白质的功能。不过半胱次磺酸不稳定,能被硫氧还蛋白还原酶和谷氧还蛋白还原成半胱氨酸。巯基阴离子被过氧化氢氧化为半胱次磺酸属于可逆过程,这符合信号调节的最常见模式。根据估计,细胞内过氧化氢浓度达到nM水平时,巯基阴离子会发生氧化。随着过氧化氢浓度的增加,半胱次磺酸可被进一步氧化,形成半胱亚磺酸或半胱硫磺酸。和半胱次磺酸不同,半胱亚磺酸或半胱硫磺酸不能被还原,蛋白质会发生不可逆永久性损伤。因此,细胞内过氧化氢浓度必须维持在一定的正常范围,才能保持细胞免受氧化损伤。细胞内维持氧化还原平衡主要依靠还原型谷胱甘肽/氧化型谷胱甘肽和NADPH/NADP的比例,而这些氧化还原对比例的维持最终依靠能量物质代谢过程中提供的电子。因此一旦细胞能量代谢受阻,细胞内氧化还原平衡必然发生紊乱,就非常容易发生氧化损伤。
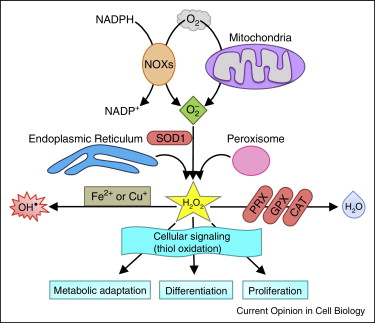
细胞避免活性氧过度增加导致氧化损伤的另一重要方式是将活性氧限制在特定部位,这样能让活性氧发挥作用,又避免对周围其他成分产生影响。例如某蛋白受来自NADPH氧化酶来源的过氧化氢调节,这种蛋白就定位于细胞膜上。SOD2(线粒体)基因敲除动物和SOD1(细胞浆)基因敲除的病理表现有很大差异。是因为线粒体内存在大量铁硫蛋白,导致线粒体内超氧阴离子聚集和细胞浆超氧阴离子聚集的作用有很大区别。总之,活性氧的种类和分布对其生物学效应影响很大。
多细胞生物利用生长因子整合细胞分裂、存活和营养摄取信号,调节细胞分裂。生长因子如表皮细胞生长因子和血小板源生长因子能依靠受体酪氨酸激酶方式,可使受体细胞内段特定酪氨酸残基自身磷酸化,随后受体能招募多种蛋白,启动多种信号传导通路。其中PI3K/AKT和RAS/MEK/ERK MAPK通路激活可促进细胞分裂、营养摄取和细胞存活。受体酪氨酸激酶和PI3K也可分别受到蛋白酪氨酸磷酸酶和磷酸酶PTEN负向调节(去磷酸化),停止细胞分裂信号。
许多生长因子信号通路通过磷酸化启动,通过去磷酸化关闭,也可以通过停止去磷酸化维持启动。活性氧在调节磷酸化和去磷酸化过程中也具有重要作用。首先,生长因子如PDGF和 EGF能快速提高活性氧的产生,主要是通过NADPH氧化酶途径,活性氧是生长因子诱导酪氨酸残基自身磷酸化的重要条件。
研究发现,生长因子诱导过氧化氢产生,后者能将蛋白酪氨酸磷酸酶1B(PTP1B)催化活性半胱氨酸氧化成为次磺酸,导致PTP1B酶活性丧失。PTP1B能将EGF受体酪氨酸去磷酸化失活,PTP1B酶活性丧失则能维持EGF受体酪氨酸磷酸化激活,使该信号途径维持发挥作用。氧化失去活性的PTP1B可以被硫氧化还原蛋白重新还原恢复活性,使这种氧化激活的EGF受体酪氨酸磷酸化失去活性,形成氧化还原调节信号的完整回路。细胞内MAPK信号是PDGF激活后的信号传导系统,这一系统的PDGF受体相关的SHP-2磷酸化也受到氧化还原的调节。
过氧化氢能体外可逆性氧化许多PTP蛋白,导致这些蛋白失去活性。人PTEN蛋白Cys121和Cys71可被过氧化氢氧化,形成二硫键而失活,这种变化可以被硫氧还蛋白还原恢复活性。细胞经各种和PI3K激活相关的生长因子短暂刺激后,氧化型PTEN增加。正常情况下,过氧化物酶可快速抑制生长因子刺激引起的过氧化氢水平升高,最近数据发现,生长因子刺激可造成细胞膜局部过氧化物酶1(PRX1)本身可被磷酸化失活,这能促进局部过氧化氢水平升高,抑制磷酸酶活性。因为PRX1失活只局限于细胞膜,细胞内PRX1仍保持活性,这样可以避免细胞内发生过氧化。这些研究说明,生长因子激活必需伴随细胞膜局部活性氧的快速一过性增加,活性氧使磷酸酶失活,使磷酸化细胞增殖信号强化。最近研究显示,除NADPH氧化酶以外,线粒体来源的活性氧也参与磷酸酶氧化失活效应。
恶性肿瘤细胞往往处于高代谢状态,活性氧水平是超过正常细胞。为维持快速增殖的状态,肿瘤细胞必须让生长因子通路处于持续激活状态,这种状态使之能吸收更多营养、存活和分裂信号增强。这种状态也必然导致线粒体、内质网和NADPH氧化酶产生更多活性氧。
20年前,科学家发现肿瘤细胞比相应正常组织细胞的活性氧水平高。推测活性氧是导致基因组不稳定,促进肿瘤发生的原因。但是随着对肿瘤发生研究的深入,这种观点逐渐受到挑战,发现活性氧促进肿瘤发生主要是通过信号调节而非染色体结构的破坏。
染色体稳定性可能主要是p53蛋白的功能不足造成的异倍体,但p53蛋白缺陷并是因为活性氧增加。p53蛋白由393个氨基酸组成。DNA损伤时,细胞主要反应之一便是p53蛋白增加。P53蛋白主要集中于核仁区,能与DNA特异结合,活性受磷酸化调控。正常P53的生物功能好似“基因组卫士”,在G1期检查DNA损伤点,监视基因组的完整性。
而依赖于活性氧的myc基因促进肿瘤的发生并不需要基因组完整性的破坏。myc基因是较早发现的一组癌基因,包括C- myc、N-myc和L-myc ,分别定位于8号染色体、2号染色体和1号染色体。myc基因属于编码核蛋白的癌基因,3个基因都编码一种与细胞周期调控有关的核内DNA结合蛋白。myc基因家族及其产物可促进细胞增殖,永生化,去分化和转化等,在多种肿瘤形成过程中处于重要地位。
重要的是,用抗氧化药物如NAC或NADPH氧化酶抑制剂减少细胞内活性氧水平可以抑制导致成纤维细胞转化的促有丝分裂信号。KRAS导致的人癌细胞分裂也需要线粒体来源的活性氧。线粒体突变时三羧酸循环或电子传递链功能紊乱产生活性氧的能激活肿瘤形成信号,如PI3K和MAPK。
活性氧的另一个目标是转录因子NF-kB。NF-kB控制肿瘤细胞的存活信号,是最早被发现可受到活性氧调节的转录因子。癌细胞内存在活性氧产生增加,抗氧化活性也高于正常细胞,达到一种动态平衡。如果癌细胞不能控制活性氧,氧化损伤同样可以导致癌细胞死亡。不同情况下,癌细胞内整体水平活性氧有可能高于正常细胞,也有可能低于正常细胞。不过在活性氧产生部位,往往因为产生速度快而引起信号效应。因此,癌细胞的特点是一方面自身抗氧化能力很强,另一方面活性氧信号作用比较强。这样的结果是在保持氧化信号很高的同时,而不会因为氧化损伤导致细胞死亡。
维持高水平抗氧化能力的关键是活性氧信号能激活抗氧化转录因子NRF2。NRF2正常情况下与KEAP1结合,后者能引导NRF2进入蛋白水解过程,导致NRF2被降解。活性氧能氧化KEAP1的氧化敏感半胱氨酸,导致NRF2与KEAP1分离,游离的NRF2进入细胞核与小MAF蛋白形成异二聚体,结合在DNA抗氧化反应序列(AREs)上,发挥转录活性,促进多种抗氧化基因的表达。除活性氧外,一些信号分子如ERK 、MAP K和PI3K也能激活NRF2。某些肿瘤细胞是因为KEAP1基因突变,导致NRF2被持续性激活。NRF2失活能使肿瘤细胞发生氧化应激,避免肿瘤发生。如果NRF2失活会导致多种抗氧化系统失去功能,导致大量活性氧的产生,破坏肿瘤细胞。但是,失去特异性抗氧化系统不仅可以导致组织损伤,本身也可以造成肿瘤的发生。
如果活性氧是促进分裂增殖信号的必要条件,通过抗氧化剂将活性氧水平降低到无法刺激分裂信号程度,应该可以抑制肿瘤的发生。肿瘤抑制因子p53可控制多种抗氧化基因表达,p53缺陷小鼠肿瘤模型可以用NAC抑制肿瘤发生提示p53控制某些肿瘤发生是通过提高细胞抗氧化能力。最近研究发现,p53控制肿瘤发生主要是促进调节抗氧化和代谢能力而不是通过促进细胞凋亡和细胞周期。p53促进抗氧化是通过TIGAR调节代谢实现的。TIGAR具有果糖2,6二磷酸酶活性,可降低果糖2,6二磷酸水平,正向调节磷酸果糖激酶1活性。这些作用能抑制糖酵解和促进通过葡萄糖磷酸戊糖途径产生NADPH,NADPH是维持抗氧化的重要介质。其他一些肿瘤抑制因子如FOXO也具有促进抗氧化系统的作用。
低氧条件下活性氧也会增加,低氧本身和活性氧可以促进HIF水平的升高,导致血管增生等低氧相关效应。
总之,活性氧和肿瘤的关系目前仍存在争议,但许多学者支持肿瘤细胞一方面大量产生活性氧,启动分裂增殖、存活和代谢适应信号。另一方面,肿瘤细胞通过提高抗氧化能力控制和压制活性氧水平,避免因严重氧化应激导致细胞死亡。
这一观点会导致通过调节活性氧治疗肿瘤产生一种尴尬,因为一种策略是通过进一步压制活性氧水平,控制因活性氧启动的促肿瘤信号,另一策略是提高细胞活性氧水平以杀死肿瘤细胞。通过系统回顾使用b胡萝卜素、维生素A和维生素C等抗氧化物质治疗肿瘤的研究发现,这些抗氧化物质对治疗肿瘤并没有帮助,甚至会产生相反效果。或许应该采用亚细胞结构的特异性抗氧化策略,如线粒体和NADPH氧化酶,以控制这些能产生启动肿瘤发生信号的活性氧,才能产生预期目的。关于这方面的临床研究目前还没有完成。许多证据表明,通过一些小分子诱导剂能通过破坏抗氧化系统提高活性氧选择性诱导肿瘤细胞死亡。不过仍有两个疑问需要克服,一是如果活性氧水平不够高,不仅产生不了杀死肿瘤细胞的目的,甚至可能增强NF-kB、PI3K、 HIF和MAPK等肿瘤生长信号。二是这种氧化应激诱导分子没有选择性进入肿瘤细胞,而进入正常细胞,导致正常细胞被破坏。因为不同类型的肿瘤细胞对氧化和抗氧化的敏感性不同,只能继续对这两种相反的策略进行更深入研究,希望能确定出有希望的协助其他治疗方法的促氧化和抗氧化新方法。
关注医学科技

https://wap.sciencenet.cn/blog-41174-887237.html
上一篇:长期饮氢水对高血压病引起的血脑屏障破坏的预防性研究
下一篇:《科学》中国科学家解释衰老分子机理