博文
美国农业部Alan J. Franzluebbers等发表美国东南部草地与农田土壤健康与根区富集特征研究论文
||

文 章 信 息

期刊名称:Grassland Research(草地研究)
英文标题:Soil health and root-zone enrichment characteristics between paired grassland and cropland fields in the southeastern United States
中文标题:美国东南部草地与农田土壤健康与根区富集特征
第一作者/通讯作者:
Alan J. Franzluebbers(美国农业部(USDA)农业研究局(ARS))
编译者: 尤 杨 兰州大学草地农业科技学院 在读博士生
说明:该文仅代表编译者对论文的理解,如需参考和引用相关内容,请查阅原文。
摘 要
研究背景——美国东南部私人经营牧场土壤有机碳和氮数据相对较为稀缺。
研究方法——在美国东南部的三个主要土地资源区,采用成对农场方法来研究拥有放牧、植物多样性的多年生牧场与一年生单一作物耕地之间的各种土壤健康参数、养分和水循环变化。
研究结果——农田土壤稳定性指数平均为0.64 mm mm−1,放牧土壤稳定性指数平均为0.91 mm mm−1,表明放牧土壤表面更稳定,具有抗侵蚀和快速入渗的能力。表层土壤有机C和N组分(即0−10 cm深度的总组分、颗粒组分和矿化组分)在放牧条件下大于耕地处理。不同地点的根区有机C和N组分(0–30 cm土层)在草地土壤的富集程度高于农田土壤。Blue Ridge(2.87 vs. 1.10 Mg N ha−1)和Piedmont(2.80 vs. 2.10 Mg N ha−1)的牧场土壤中根区土壤氮的富集程度高于农田(p < 0.05),但在Blackland Prairie(2.40 vs. 2.12 Mg N ha−1)两者差异不显著。
研究结论——本研究表明,与邻近生产商品饲料粮的农田相比,多年生轮牧草地可以储存更多的土壤有机碳和氮,并改善土壤表面稳定性。
前 言
以牧场为基础的畜牧业生态系统服务尚未被广泛描述。然而,越来越多的证据表明,多年生草地能够提供多样的生态系统服务,有助于保护农业生产和农业赖以生存的环境(Lemaire et al., 2011; Sollenberger et al., 2019)。有证据表明,在温暖湿润地区,例如美国东南部的牧场管理下,土壤有机碳积累显著增加(Barral et al., 2007; Franzluebbers, 2010)。对于这种表层土壤有机碳的变化如何影响其他土壤性质,例如水分入渗和储存、表层土壤团聚体以及具有生物活性的C和N组分,目前尚未引起足够重视。团聚体和生物活性有机C、N组分的变化也可能是导致土壤微生物群落结构发生变化的重要步骤(Yu et al., 2021)。一个地区的不同农场需要根据土地利用的长期变化确定土壤属性,因为农场管理和固有土壤属性可能对生态系统服务结果产生巨大影响(Franzluebbers & Poore, 2021; Kaiser et al., 2016)。
最近的证据表明,土壤有机质的生物活性组分可能是改变不同土壤条件下与养分和水分循环有关的表层土壤特征的最重要因素(Franzluebbers, 2016, 2022a)。因此,本研究旨在获取农田数据,以验证或反驳以往研究站的结论,即草地具有显著的土壤有机碳固存和土壤聚集特征,并探索私营农场不同土地利用方式下土壤健康状况的一般特征。
牛肉提供人类必需的营养物质,如蛋白质、锌、铁和B12(Agarwal & Fulgoni,2022)。此外,相较于谷物喂养的牛肉,草食喂养的牛肉具有更高比例的3: omega 6脂肪酸和更多共轭亚油酸,从而提供了潜在的对人类健康有益处的物质(Daley et al., 2010)。在牧场上吃各种各样的植物的牛也获得了多种具有潜在抗炎、抗癌和心脏保护作用的植物化学物质和生物化学物质,这些物质集中在肉类中:萜类、酚类、类胡萝卜素、抗氧化剂、肽和其他生物活性化合物(Van Vliet et al., 2021)。我们假设,与传统的谷物混合饲料相比,在营养丰富的牧场土壤上放牧的牛采食多种牧草可能会拥有更为丰富的肉类植物化学物质。基于土壤-植物-牛-人类健康连续体的假设,我们计划进行一项研究,旨在探索通过土壤健康状况差异来引导以饲草为主要成分的牛饲料,与传统混合玉米饲料喂养肉牛系统相比的可行性。具体来说,本研究的这一部分旨在描述放牧、植物多样性牧场和邻近生产用于饲育的商品饲料谷物的农田之间土壤特性的差异。
材料和方法
从2021年5月6日至18日,在3个地点以500 ± 240 km为间隔,对草地和农田进行了成对采样。在北卡罗来纳州(南Blue Ridge主要土地资源区[MLRA];35.50° N, 82.37° W)的Buncombe县的一个农场中,选取了1 km范围内的4个不同的牧场,并与距离12 km的农田中的4个样本进行匹配Ashville,NC年平均气温(1991–2020)为14.1°C,年平均降水量为1030 mm(美国国家海洋和大气管理局[NOAA], 2023)。同样,在北卡罗来纳州Surry县的单个牧场((南Piedmont MLRA;36.85° N, 79.48° W)上采样距离1 km范围内的4个牧场,并与邻近农户农田田块(<2 km间隔))的4个样本进行匹配。年平均气温在Mt. Airy的平均气温为13.1°C,年平均降水量为1280 mm。最后,在阿拉巴马州Perry县(阿拉巴马州和密西西比州黑土草原MLRA;32.56° N, 87.45° W)的一个农场上采样了3 公里范围内的4个牧场,并与距离3.4 公里的邻近农户农田中的4个样本进行匹配。阿拉巴马州Greensboro年平均气温为17.8°C,年平均降水量为1392 mm。
牧场由多种牧草混合组成,包括位于Blue Ridge的高羊茅(Lolium arundinaceum [Schreb.] S.J. Darbyshire Lolium arundinaceum [Schreb.] S.J. Darbyshire)、鸭茅(Dactylis glomerata L.)、车前草(Plantago lanceolata L.)和白三叶(Trifolium repens L.),位于Piedmont的高羊茅、红三叶(Trifolium pratense L.)、鸭茅和马唐(Digitaria sanguinalis [L.] Scop.),以及位于Blackland Prairie的高羊茅、黑麦草、车前草和菊苣(Cichorium intybus L. Cichorium intybus L.)。所有农田均种植玉米(玉米Zea mays L.),在Piedmont和Blue Ridge地区进行保护性耕作,在转化土草原地区进行传统耕作。在Blackland Prairie地区采用常规耕作。牧场和农田各自的土地用途至少有10年。
每个畜牧农场都会精心选择土壤和景观位置,以与附近用于生产牲畜饲料谷物的农田相协调。我们可以预料到这些地点的一些差异,因为在过去几十年持续使用土地之前的事件并不为人所知。然而,在本研究中,相似的土壤类型的假设并不重要,因为我们通过根区富集计算程序考虑了亚土壤C和N浓度的差异(Franzluebbers, 2021b, 2023)。土壤描述来自美国农业部土壤调查地图。在Blue Ridge地区,土壤为Dillard壤土,坡度1%−5%,很少被淹(细壤土、混合壤土、次活跃壤土、中间潮土)和Iotla壤土,坡度0−2%,偶尔被淹(粗壤土、混合壤土、活跃壤土、中间潮土)。在Blue Ridge地区的4个牧场中,土壤类型包括Evard-Cowee复合体、盆地和8%-15%斜坡上的石质土壤(细壤型、副土型和中间型典型单坡型),Toecane-Tusquitee复合体、15%–30%斜坡上的巨砾土壤(壤土骨架型、混合型、活跃型和中间型腐殖质单坡型),Tate壤土,盆地和15%斜坡上的不同类型土壤(细壤型、混合型、半活跃性和中间性典范单坡形式)以及Dillard壤土,在1%-5%斜坡上。在山前地区,农田和牧场具有相似的土壤系列,主要为Fairview砂质粘性黏土,并分布于2%-8%和8%-15%的坡度之间,存在着适度侵蚀现象(细高岭泥矿物质介素类)。此外还有一种被称为Colvard和Suches的0-3%坡度下偶尔发生淹水情况下出现的特殊类型(粗壤型,混合型,活性,非酸性,中间型)。在Blackland Prairie,牧场位于0−3%坡度的Okolona粉质粘土壤土(细粒、膨润土质、热性的氧化水稻土)和1%−3%和3%−8%坡度的Sumter粉质粘土壤土(中度侵蚀)上(细粉质、碳酸盐、热溶质沉积物),农田位于1%−3%坡度的Vaiden粘土(非常精细的,微晶的,热流体结构)上。每个位置的单一农田的重复样本以>30 m的距离分开。
采用内径为4 cm的推入式土钻在0−10 cm深度进行土壤采样。对于每个重复,我们合成了5个子样本(在主要方向上与中心点相隔10 m)。一旦收集到0-10 cm的样品,用电池供电的螺旋钻(直径3.2 cm)在7.6 L桶的底部通过一个钢加固的孔钻到30 cm的深度。在5个孔中的3个,用钻柱进一步挖掘30-60 cm深的土壤。在烘箱中55℃下将土壤干燥3天。记录干重并用于计算0-10 cm深度下已知体积(628 cm3)的容重。在0-10cm深度处,干燥的复合样品重量为756±52 g,在10-30 cm深度处为487±152 g,在30-60 cm深度处为588±159 g。土壤经过4.75 mm筛网过筛。经过筛选,土壤密度通过使用金属铲子(59-和79-mL)刮取到一定体积后的平均质量来确定。该方法与Franzluebbers(2021a, 2022b)的研究报告相同。
在1-L圆柱体中,将59 mL土壤加入80 mL浓度为0.1 mol L−1的Na4P2O7溶液中,并进行16小时振荡。随后,在沉降5 h时使用比重计对土壤质地进行测定。随后,通过将悬浮液经由具有0.053 mm开口的筛网过滤,并转移到干燥瓶中以测定其干质量来进行砂的测定。泥沙浓度计算为这两个组分的差值。颗粒有机碳和氮浓度通过干燥和球磨砂进行测定。非颗粒有机碳和氮含量来源于土壤中有机碳与颗粒有机碳、以及土壤全氮与颗粒有机氮之间的差异。这种组分有时被称为矿物结合的有机碳和氮,但尚不清楚这种组分是否只包括矿物结合的碳和氮,还是也包括溶解的和松散的结合的碳和氮,这些结合的碳和氮没有大到可以捕获成颗粒大小,即>0.053 mm。因此,这里称之为非颗粒有机C和N。
根据北卡罗来纳州农业土壤测试服务的常规程序,分析了土壤化学性质。土壤测试生物活性(STBA)是指在将干燥的土壤(59 mL)重新湿润至50%的水填充孔隙空间后,在25°C下孵育3天期间释放出的二氧化碳量(Franzluebbers, 2022a)。在土壤体积恒定、质量可变的情况下,根据59 mL土的筛分土壤密度,调整各样品的加水量,使孔隙水占50%(孔隙气占50%)。在相同条件下,通过进一步培养土壤至24天来测定累积碳矿化。基础土壤呼吸是培养3−24天碳矿化的假定线性速率。用球磨机将20-30 g子样品磨成细粉,然后用Leco TruMac(Leco Corporation)燃烧0.5 g子样品,测定土壤总碳和氮。除Blackland Prairie的两个牧场外,我们假定所有样本的土壤总碳代表有机碳。无机C通过3 mol L−1 HCl溶解碳酸盐后土壤质量的差异来确定。
土壤团聚体在北卡罗莱纳州立大学土壤生态与管理实验室使用标准化程序进行测定(Franzluebbers, 2022a, 2022b)。在开口为1.0、0.25和0.053 mm的筛网上摇晃1 min,从79 mL体积的土壤中分离出干稳定性团聚体。在直接浸入水中并振荡10分钟(20 mm冲程,30次/min)后,将水稳定的团聚体隔离在开口为1.0和0.25 mm的筛网上。将经过0.25 mm筛子的含水土壤倒在0.053 mm筛子上。<0.053 mm组分作为初始土壤质量与其他组分总和的差值。所有组分在55°C下烘干24小时以达到恒定质量。干稳性和水稳性团聚体的平均重量直径分别由比例分数和平均直径的乘积计算。土壤稳定性指数计算为水稳性团聚体的平均重径与干稳性团聚体的平均重径之比。
根据Franzluebbers(2021b, 2023)先前的描述,根据三个采样深度的浓度和容重的土壤传递函数(BD = 1.71 e[−0.013×SOC])计算土壤有机C和N组分的根区富集程度。作为对每个有机C和N组分重复的例子,土壤有机C的浓度拟合为依赖于土壤深度的非线性函数,根据:
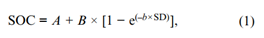
其中,SOC为土壤有机C(g kg−1),A为不受管理影响的剖面深层C浓度(A≥0),B为土壤表层积累的有机C库,并随土壤深度(SD,|cm|)呈指数函数下降。所有非线性回归都使用SigmaPlot v. 14(Systat Software Inc.)拟合可用数据。如果方程产生A < 0的估计值,则使用修改后的形式将A设为0:
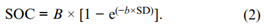
当浓度乘以区间上限和下限的预测容重值时,利用方程计算出增量为5 cm深度的有机C和N组分浓度,以及5 cm至30 cm深度的有机C和N组分含量。这些增量的总和表示了地表30 cm剖面内C和N组分的储量。因此,所有三个采样深度(0–10 cm、10–30 cm和30–60 cm)被用来通过回归对每个领域/重复产生一个单一的总含量估计值。不受管理影响的基线有机碳和氮组分被假定为30 cm深度的浓度(Franzluebbers,2021b)。基线有机C和N组分含量(0–30 cm深度)是由30 cm深度的浓度乘以30 cm深度的估计容重得出的,该结果预测了整个0–30 cm剖面。根区有机碳和氮组分的富集是根据每个剖面的总储量和基线储量之间的差异来计算的。
使用SAS(SAS Institute Inc.)的一般线性模型对(a)地点内的变化进行分析,其中包括由于复制的随机变化和土地利用的固定变化所导致的差异;同时,还对(b)地点间的变化进行分析,其中包括由于复制的随机变化、位置和土地利用以及位置×土地利用交互作用所导致的差异。这些差异在统计学上具有显著意义(α=0.05)。
结 果
土壤物理性质
Blue Ridge、Piedmont和Blackland Prairie的三个地点提供了土壤质地和气候条件的多样性,以比较放牧牧场和农田对土壤性质的影响。所有三个地点和采样深度(0–10、10–30和30–60 cm)的土壤质地成分的范围如下:粘土为185−602 g kg−1(5%−95%范围),粉砂为154−405 g kg−1,沙子为49−612 g kg−1。在Blue Ridge和Piedmont地区,作物处理和牧场处理在土壤质地上只有少数显著差异,且仅在0−10 cm深度,而不是更深,这表明土地利用在同一地点的类似土壤类型上是合理匹配的。Blue Ridge区草地土壤粘土含量较低(166 vs. 261 g kg−1),淤泥浓度较高(286 vs. 212 g kg−1)。在Piedmont地区,放牧条件下的粉土比例(199 vs. 159 g kg−1)也大于农田条件下的粉土比例。在所有四个重复中,土壤质地组分别是Blue Ridge地区作物和牧场下的砂质粘土壤土,Piedmont地区两种土地利用下的砂质粘土壤土,Blackland Prairie地区作物和牧场下的粉质粘土和粘土。在不同地点,在0−10 cm深度,草地下的粘土含量比农田下的低,淤泥含量比农田下的高(表1),这可能是由于不同农田之间的历史侵蚀差异造成的。在10−30 cm和30−60 cm深度,草地下的粉土比例大于农田下。这些成对农场土地利用之间土壤质地的细微差异使我们有理由探索根区富集计算的概念,以将土壤控制因素对有机质组分的影响与管理/历史土地利用的影响分开。
草地的土壤筛分密度低于农田,但仅在0–10 cm深度(表1)。0−10 cm土层的土壤容重在不同土地利用方式间无显著差异(表2)。样品初加工去除的石质组分(>4.75 mm)在不同土地利用方式之间也没有差异。因此,这两种土地利用方式对土壤物理状况的典型指标的影响并不一致。
表1. 美国东南部三个成对农田和放牧牧场在三个深度的土壤物理和生物地球化学特性
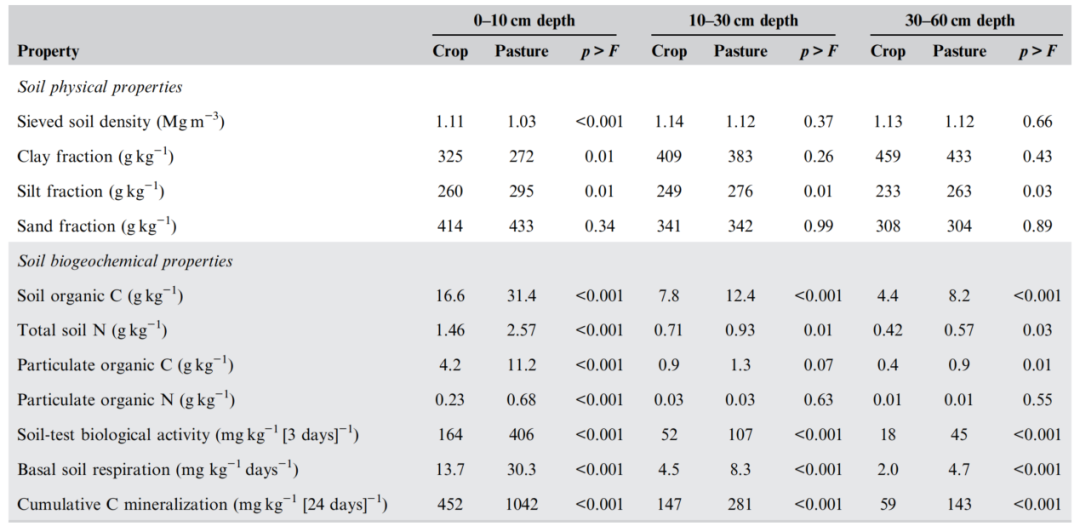
表2. 在美国东南部的3个地点,在0−10 cm深度成对的农田和放牧草场下的土壤物理和化学性质
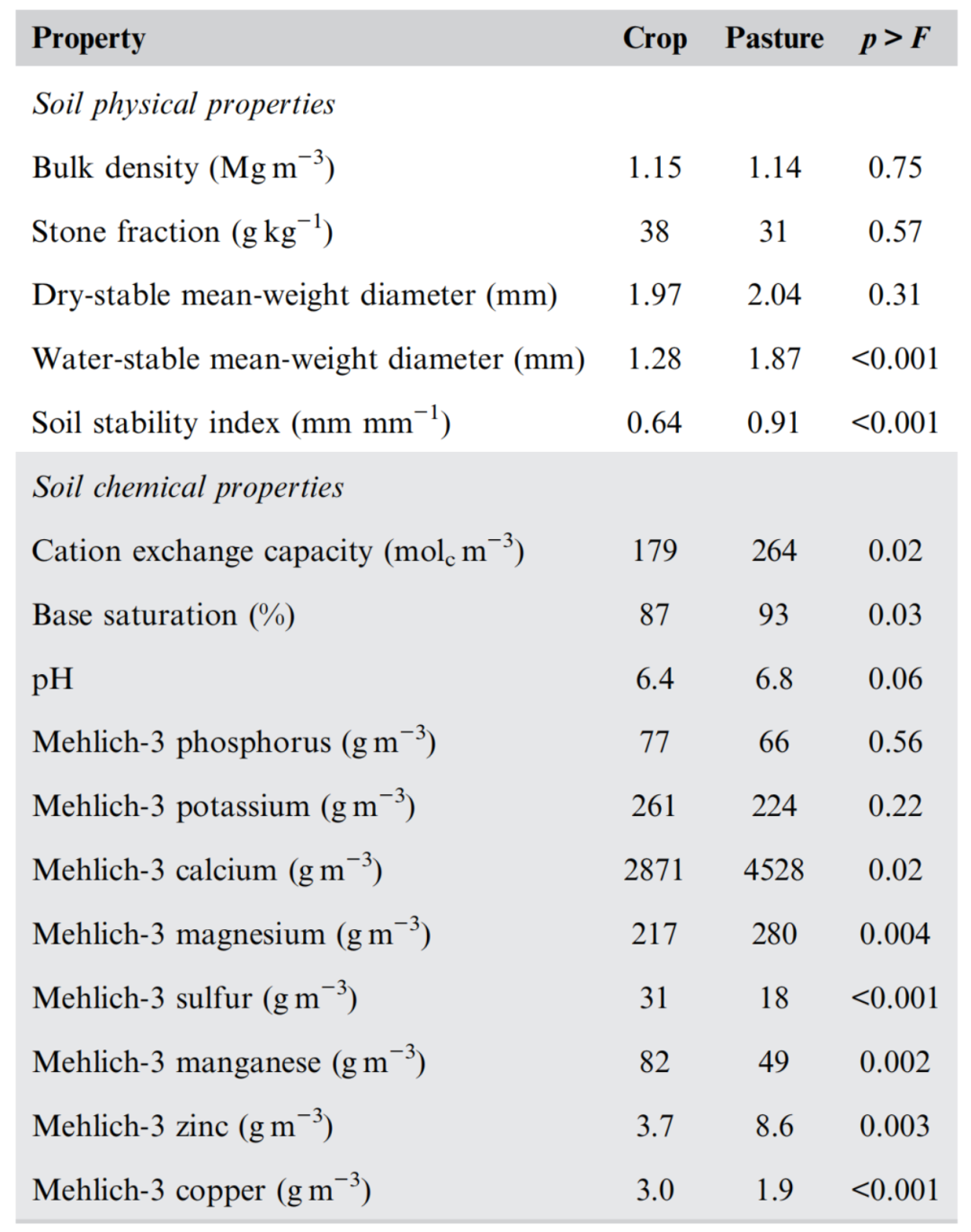
土壤团聚体的干稳定平均重量直径在不同土地利用方式之间没有差异(表2)。尽管在0-10 cm深度(85-615 g 沙土 kg−1;5%-95%范围),干稳定平均重径仅在1.7–2.4 mm之间变化。水稳定性平均重径在0.8−2.3 mm之间变化,且草地下的水稳定性平均重量直径显著大于农田下。Blue Ridge、Piedmont和Blackland Prairie的水稳性平均重量直径分别为0.86和1.73 mm(p < 0.001)、1.27和1.70 mm(p < 0.001)和1.72和2.18 mm(p < 0.001)。草地土壤稳定性指数(水稳定性除以干稳性平均重径)也大于农田土壤。这种效应在每个单独的位置也很显著。水稳定性平均重量直径和土壤稳定性指数与STBA密切相关(图1)
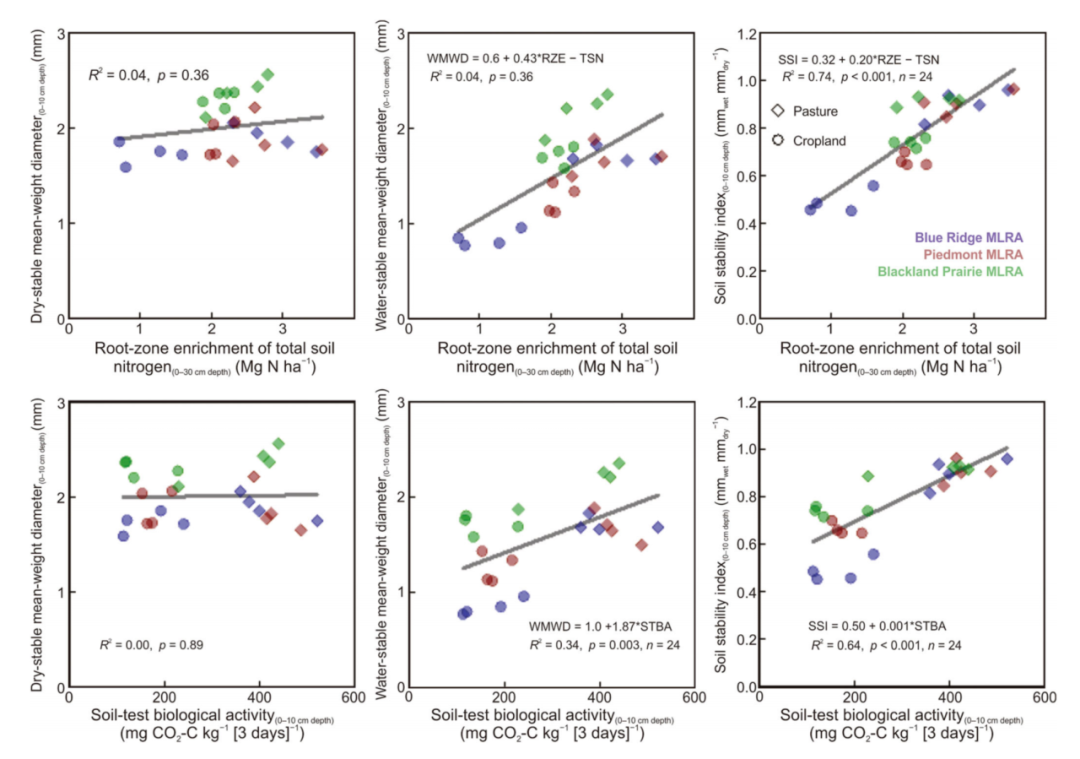
图1. 北卡罗莱纳州(Blue Ridge和Piedmont主要土地资源区[MLRA])和阿拉巴马州(Blackland PrairieMLRA)三个地点不同土地利用方式下,干稳定平均重径、水稳定平均重径和土壤稳定性指数与根区总氮(0-30 cm深度)富集和土壤试验生物活性(0-10 cm深度)的关系
土壤化学性质
在不同地点,0-10 cm深度的土壤化学性质受土地利用方式的影响(表2),但pH(6.6 ±0.8 )、Mehlich-3可萃取P(71 ± 59 g m-3)、Mehlich-3可萃取K(243 ± 83 g m-3)和Mehlich-3可萃取Mg(249 ± 120 g m-3)不受土地利用方式的影响。放牧草地的阳离子交换量、盐基饱和度、Mehlich-3可萃取Ca和Zn含量均高于农田。草地土壤Mehlich-3可萃取S、Mn和Cu含量低于农田。Blackland Prairie的两个牧场含有影响总土壤C的碳酸盐。从这两个牧场的土壤深度计算,碳酸盐-C为18.0±2.2 g kg−1。
在粘土富集的Blackland Prairie地区,阳离子交换容量(457±167 molc m−3)明显大于其他两个地区(104±38 molc m−3),这可能是由于粘土含量高,以及粘土类型(2:1的晶格结构,而其他地区为1:1)。影响阳离子交换能力的另一个因素是草地土壤有机碳和全氮含量高于农田土壤(表1和表2)。Piedmont地区的基础饱和度(79%±13%)低于其他两个地区(96%±4%),这可>能是由于林地清理时间较近(<15年),而其他地区较近(>50年)。Piedmont地区的土壤pH值(5.9±0.7)也低于其他两个地区(6.9±0.6),部分原因是由于不同地区可用于改善酸度的时间不同以及土壤形成因素的差异。Blue Ridge地区Mehlich-3可提取磷含量(101±76 g m−3)高于Blackland Prairie地区(30±23 g m−3),Piedmont地区为中等水平(83±42 g m−3)。Blackland Prairie的Mehlich-3可提取钾含量(298±103 g m−3)高于Piedmont的195±19 g m−3,Blue Ridge的Mehlich-3可提取钾含量为234±72 g m−3。Blackland Prairie的Mehlich-3可提取钙含量(8346±3633 g m−3)高于其他两个地点(1376±704 g m−3)。Blackland Prairie地区的可提取Mg含量(299±111 g m−3)高于Piedmont地区(183±127 g m−3),Blue Ridge地区为中等水平(264±92 g m−3)。Piedmont的Mehlich-3可萃取S(32±15 g m−3)大于其他两个区域(21±7 g m−3)。Mehlich-3可提取Mn的顺序为:Blue Ridge(103±50 g m−3);Blackland Prairie(67±35 g m−3);Piedmont(27±18 g m−3)。Mehlich-3可萃取Zn在Piedmont(10±8 g m−3)大于其他两个区域(4±3 g m−3)。不同区域的Mehlich-3可萃取铜含量无差异。
土壤生物地球化学性质
土壤有机碳在0-10 cm深度(24±10 g kg−1)最大,在10–30 cm深度(10±5 g kg−1)下降一半以上,在30-60 cm深度(6±5 kg−1)进一步下降较小。土壤全氮浓度在0−10 cm深度为2.0±0.8 g kg−1,10−30 cm深度为0.8±0.4 g kg−1,30−60 cm深度为0.5±0.3 g kg−1。土壤有机碳和全氮浓度在这些深度增量内的主要差异是由土地利用/管理造成的(表1)。在所有三个深度,牧场土壤有机碳和全氮浓度都高于农田土壤。
在0−10 cm深度,Blackland Prairie土壤有机碳(29.6±12.1 g kg−1)高于其他两个地点(21.2±7.6 g kg−1)。这一差异在较低深度也一致(Blackland Prairie在10-30 cm深度为14.7±7.0 g kg−1,其他两个地点为7.8±1.9 g kg−1);在30-60 cm深度分别为10.4±6.2和4.2±1.8 g kg−1)。在0−10 cm深度和10−30 cm深度,Blackland Prairie土壤全氮(2.45±0.90 g kg−1)大于其他两个位置(1.80±0.63 g kg−1)(分别为1.08±0.43和0.69±0.22 g kg−1)。Blackland Prairie 30−60 cm土层全氮(0.66±0.31 g kg−1)高于Piedmont(0.30±0.11 g kg−1),Blue Ridge处于中间水平(0.52±0.14 g kg−1)。
在每个位置(数据未显示),在0-10 cm深度的所有位置(表1),放牧草地下的颗粒有机C和N大于农田下。在较低深度,放牧草地下的颗粒有机C趋势较大(10-30 cm),而放牧草地下的颗粒有机C趋势较大(30-60 cm)。土壤深层颗粒有机氮受土地利用方式影响不显著。
在不同地点(表1),以及在每个地点(数据未显示),放牧草地下24天内的STBA、基础土壤呼吸和累积碳矿化在统计学上大于农田下(表1)。与总有机碳和颗粒有机氮组分相比,这些生物活性组分的显著差异在整个采样剖面中持续存在。
根区土壤有机碳氮组分的富集
在不同地点进行分析时,放牧草地下根区土壤有机碳的富集量大于农田下(表3)。这种效应在Blue Ridge(草地和农田下分别为33.5和13.7 Mg C ha−1)和Piedmont(37.0和24.8 Mg C ha−1)有显著差异,但在Blackland Prairie地区(25.0和24.6 Mg C ha−1)没有显著差异。土地利用×区位的交互作用与土壤有机碳总储量的交互作用相反,而不考虑土壤底土条件,土壤有机碳被认为是由土壤控制的。因此,不能得出放牧比耕地普遍提高土壤有机C的结论,根区富集的计算允许更保守的估计来解释显著的底土差异,从表3中土壤有机C基线储量的显著差异中可以看出。
表3. 在美国东南部三个地点成对的农田和放牧牧场下计算0-30 cm深度的土壤属性

在不同地点的分析中,放牧下土壤全氮的根区富集也比农田大(表3)。由于海拔梯度的差异,存在显著的位置×土地利用效应,即Blue Ridge草地根区富集大于农田根区(p <0.001)(分别为2.87 vs. 1.10 Mg N ha−1),Piedmont草地根区富集大于农田根区(p =0.02)(分别为2.80 vs. 2.10 Mg N ha−1),Blackland Prairie草地根区富集与农田根区富集无差异(p =0.34)(分别为2.40 vs. 2.12 Mg N ha−1)。为了说明根区富集比土壤全氮的总库存量估计更敏感,土壤全氮的总库存量的均方根误差为0.98 Mg N ha−1,而土壤全氮的根区富集的均方根误差降低到0.39 Mg N ha−1。
与基线土壤总氮一样,颗粒有机氮的基线储量在牧场和农田之间没有显著差异(表3),这表明管理没有影响30 cm以下的颗粒有机氮,并且牧场和农田之间的匹配很好。颗粒有机碳的基线储量在放牧草地上高于农田。尽管在基线颗粒有机碳上存在这种差异,但在草地下,颗粒有机碳的根区富集程度是农田的两倍以上,而且这种效应在不同地点之间一致(如“地点×管理”不显著)。土壤中颗粒有机氮的根区富集量是农田的两倍多。
与农田相比,放牧的土地具有更大的生物活性碳库基线储量(表3)。尽管这种差异可以被认为是由于长期的土地利用对剖面更深层次的碳输入产生的影响,但也可能是由于景观位置和各种土壤形成因素的微小差异,这些差异改变了用于准备矿化的碳的生物有效性。根据基线总有机碳和有机氮储量以及颗粒有机碳和氮储量的差异,后一种解释在此处被视为更具意义。根区富集计算程序在减去30 cm根区对碳富集的影响后,独立地考虑每个土壤剖面的主根区(0-30 cm深度)碳富集的总和。即使牧场下30 cm深度的STBA基线浓度高于农田(分别为63 vs. 30 mg kg−1 [3 days]−1),上述基线条件下放牧牧场下的根区富集量仍是农田下的两倍(表3)。基础土壤呼吸(即干燥土壤复湿后CO2初始冲刷后碳矿化的线性估计)和累积碳矿化对土地利用的响应与STBA非常相似。
STBA的根区富集与被动颗粒有机碳库的根区富集和土壤全氮的总有机质库的根区富集高度相关(图2)。这些强关联独立地验证了根区富集计算对于区分0−30 cm主根区土壤成因因素和管理控制对有机质的影响的价值。此外,土壤全氮(0−30 cm深度)的根区富集与表层土壤(0−10 cm深度)的水稳性平均重量直径和土壤稳定性指数高度相关(图1)。
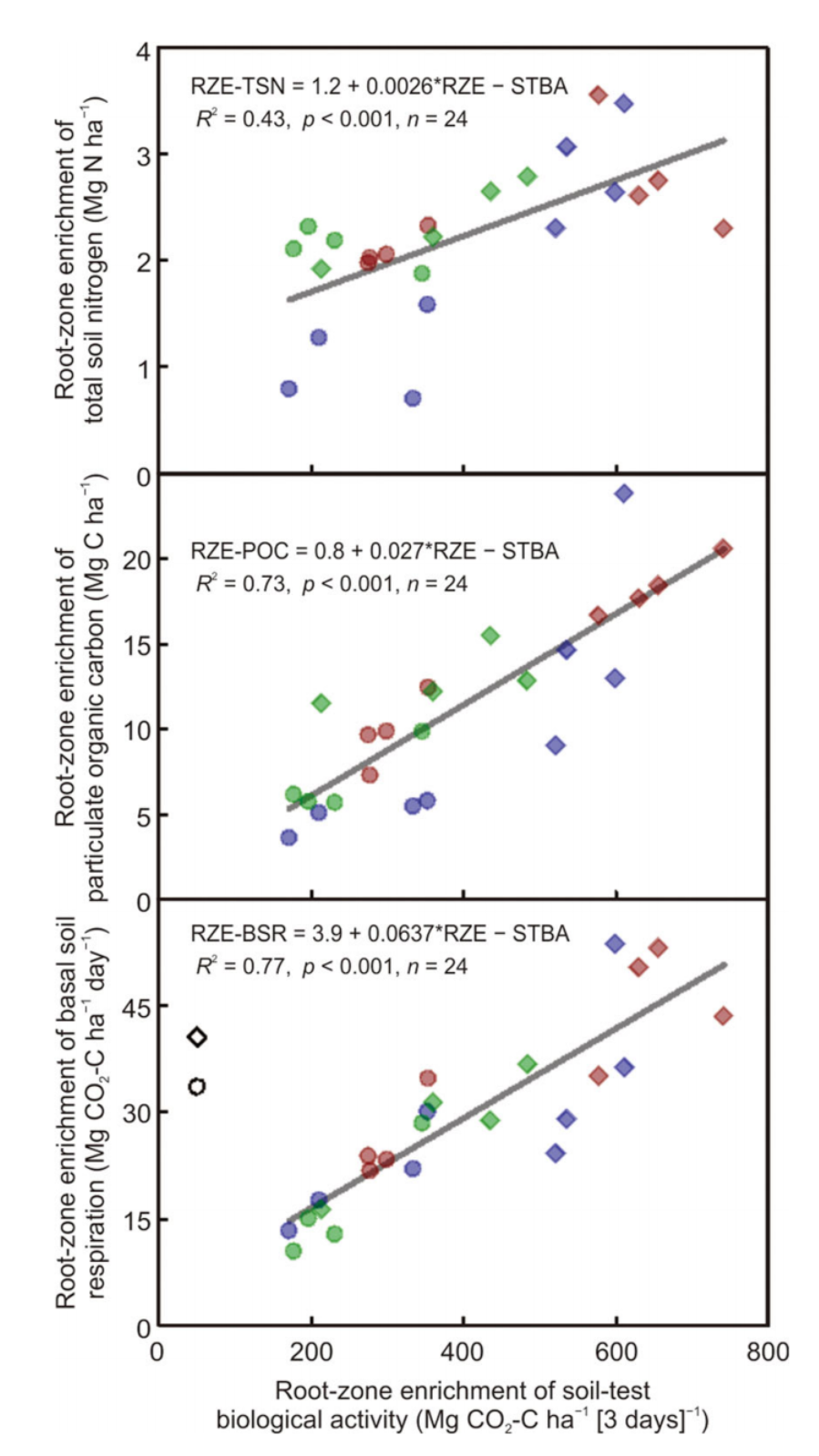
图2. 北卡罗莱纳州(Blue Ridge和Piedmont主要土地资源区[MLRA])和阿拉巴马州(Blackland PrairieMLRA)不同土地利用方式下根区土壤总氮、颗粒有机碳和基础土壤呼吸富集与土壤试验生物活性的关系
在不同地点评估时,非颗粒有机碳的根区富集在放牧牧场和农田之间没有差异(表3)。然而,土地利用×地点交互作用显著。在Blue Ridge地区,牧场根区非颗粒有机碳富集量大于耕地根区(分别为18.2 vs. 8.7 g kg−1,p = 0.004),在Piedmont地区差异不大(分别为19.7 vs. 14.8 g kg−1,p=0.11),在Blackland Prairie地区,牧场根区有机质富集量低于耕地根区(分别为8.7 vs. 17.8 g kg−1,p = 0.006)。根区非颗粒有机氮富集的响应略有不同,在Blue Ridge位置(分别为1.71 vs. 0.82 g kg−1, p < 0.001),在Piedmont位置(分别为1.66 vs. 1.53 g kg−1, p = 0.51)和Blackland Prairie位置(分别为1.62 vs. 1.73 g kg−1, p = 0.57),大于农田位置(分别为1.71 vs. 0.82 g kg–1, p <0.001)。
讨 论
在0−10 cm深度,较低的筛分土壤密度与较低的沙子浓度和/或较高的土壤有机质浓度相关(Franzluebbers, 2021a)。在测量和预测容重的24项比较中(使用基于土壤有机碳浓度的土壤传递函数),均方根误差为0.186 Mg m-3,大于在北卡罗来纳州25个研究站的研究中观察到的0.116 Mg m-3 [Franzluebbers, 2023])。
图1中土壤测试生物活性与水稳性聚集之间的关联是合乎逻辑的,因为在聚集过程中,有机质和微生物胶将土壤转化为团聚体被认为是重要的(Abiven et al., 2009),以及各种化学结合剂(Crawford et al., 1997)此前,在北卡罗来纳州的各种土地用途和地点,土壤测试的生物活性与水稳定的平均重量直径之间存在强关联(Franzluebbers, 2022b)。研究结果表明,即使农田实施保护性耕作,放牧草地的水稳性聚集性也高于农田。在这三个具有多样性土壤质地条件的地区,多年生牧草植被比一年生农田对有机质的增强生物处理(黏合土壤颗粒)更占优势。
土壤形成因素可能影响背景土壤的化学特征。然而,各种无机和有机改良剂的管理历史也可能导致不同土地用途和不同地点的土壤化学性质的差异根据开发和分析该测试的北卡罗来纳州的现场校准,Blackland Prairie地区土壤试验磷水平较低,可能会限制植物生产(Hardy et al., 2014年),但在其他州进行的类似现场校准表明,该水平可能已经足够了(Drescher et al., 2021年)。Mehlich–3可萃取K被认为适合于优化所有地点的植物生产。Mehlich–3提取的钙在所有地点都是充足的,在转化土Prairie地点的高钙区域是由于在牧场下的两个土壤中存在碳酸盐。Mehlich–3可提取态Mg、S、Mn、Zn和Cu的浓度处于植物生产所需的水平,但不会超过引起毒性的水平。
土壤有机碳和全氮随深度下降在大多数高地土壤中很常见,美国东南部土壤的典型下降幅度较大(Farmaha et al., 2022;Franzluebbers, 2021b),以及其他地区的各种土壤(Grand & Lavkulich, 2011;Hobley & Wilson, 2016),包括盐沼(Bai et al., 2016)。放牧草地下的土壤有机碳和土壤全氮浓度预计比农田下更高,因为土壤扰动较小,全年存在多年生根系,全年水分利用率更高,以及植物残留物和牲畜粪便输入的质量,特别是在地表附近(Bronson et al., 2004;Franzluebbers和Stuedemann, 2010;Franzluebbers et al., 2000)。Blackland Prairie样地土壤有机碳含量较高,可能是由于草原草地下的成土发育和粘土浓度高、阳离子交换能力强所致。
多年生牧场的地表植物残渣输入、粪便沉积和强烈的地表生根会集中在近地表的C输入,而在牧场和农田之间,随着深度的根系输入可能更相似。以前已经观察到颗粒有机碳和氮的表面变化,但很少观察到随深度的变化(Franzluebbers & Stuedemann, 2002)。
土壤试验生物活性、基础土壤呼吸和累积碳矿化反映了有机质的生物活性部分,并依赖于为土壤微生物群落提供栖息地和食物的碳输入。这些生物测量是对土壤微生物活动的群落水平估计,不能区分微生物群体,如细菌和真菌。这些措施还与净氮矿化高度相关(Pehim-Limbu & Franzluebbers, 2022),这是一个为活跃生长的植物持续提供缓慢释放的氮的过程,无论是在放牧的牧场上的多年生植物,还是在成对的谷物种植系统中的一年生植物。
特别是在进行土地利用比较和随机选择大田作为重复时,土壤控制条件可能会高度变化(a)重复之间土壤有机C和N组分存在巨大差异,限制了对处理效果的显著性检测;或者(b)声明存在显著差异时,管理以外的影响可能会干扰效果的测量。根据土壤有机碳和氮的深度分布进行根区富集计算,可以将由土壤生长因子主导的基线浓度与管理对土壤有机碳和氮浓度的影响分离开来(Franzluebbers, 2021b, 2023)。管理影响主要发生在土壤的上部30 cm,即主根区。此外,计算根区土壤有机碳和氮组分的富集,可以沿着平滑的深度过渡估算自然下降的浓度,而不是基于任意的采样深度。
我们首次估计了根区富集的除STBA以外的生物活性C库,并且这些组分的结果一致。这种一致性表明,根区富集大量土壤有机碳和氮组分可能与理解当代管理对土壤健康影响相关。
在至少10年的管理中,根区富集计算与土壤物理和生物属性的变化密切相关(图1和2)。当然,在不同的管理风格和环境下进行更多研究对于进一步发展这种土地利用对表层土壤变化影响的一致性以及在全球估算中获得更精确意义将是重要的。
非颗粒有机碳和氮在根区富集计算中,因土地利用而发生的随机变量最多且变化最小。由于非颗粒有机C和N库是高度加工和稳定的有机物部分,可能不会受到当代管理影响而发生变化(Cotrufo et al., 2019;Mosier et al., 2021)。
多年生牧草相对不受干扰的特性以及植物枯落物和粪便返回表层土壤可能是导致土壤有机碳和氮组分浓度高于农田的原因,尽管农田目前采用免耕管理。这一结果与其他研究的结果相似(Franzluebbers et al. 2000; Qiu et al., 2012)。土壤表面10 cm中总碳、颗粒和生物活性碳和氮组分的变化可能也控制了土壤团聚体的稳定性(图1)。这一观察结果验证了北卡罗来纳州其他地区土壤有机碳和氮组分与团聚体之间的关联(Franzluebbers, 2022a)。干-稳重平均直径无差异,表明不同土地利用方式下土壤团聚体的环境或化学结合机制没有改变,但在多年生牧场下,这些土壤团聚体在水中的稳定性显著增强。本研究表明,土壤有机碳和氮组分的变化体现在表层土壤物理变化中,这可能会改善农业景观中的水循环,以及减少侵蚀和养分流失。
结 论
经过对农田土壤有机碳、氮组分和水稳性聚集度的评估,本研究验证了草地畜牧业对土壤有机碳、氮积累以及与水分和养分循环相关的生态系统服务多样性以及生境发展对生物多样性的影响。通过计算根区土壤碳、氮组分的富集,揭示了这些特征在土壤剖面上的分布,进一步证实了草地管理在为生态系统服务提供养分储存、大量有机碳和氮积累以及稳定土壤表面抵御侵蚀和蓄水方面的重要性。从总量到稳定组分再到活性组分的定量分析显示出长期草地下表层土壤具备较强正向生物物理变化。
声明:该编译文章仅代表编译者对原文的理解,如需参考和引用相关内容,请查阅原文。编译文章由GR团队制作仅供学术交流,转载须注明转载自Grassland Research微信公众号及编译作者信息。
排版:甘露雪 彭京伦
统筹:沈锦慧 秦泽平


https://wap.sciencenet.cn/blog-3568995-1430716.html
上一篇:GR榜单 | 畜牧学相关微信公众号排行榜(2024年3月)
下一篇:GR榜单 | 草学、生态学、植物学及作物学等相关微信公众号排行榜(2024年3月)
全部作者的其他最新博文
- • Grassland Research在中国草学会2024年会各项活动圆满成功!感谢您的关心支持!
- • 兰州大学草地农业科技学院教师参加国家重点研发计划项目年度总结会
- • 中国科学院西北生态环境资源研究院赵鹏善研究员等:沙蓬,一种适宜中国北方沙漠和边际土地的未来作物
- • 美国佐治亚大学等机构学者:刈割策略对Bulldog 805紫花苜蓿+Tifton 85狗牙根混播草地产量及品质的影响
- • 美国堪萨斯州立大学、密苏里大学等机构学者研究成果:土壤水分管理策略和品种多样性对紫花苜蓿产量、营养品质和农场盈利能力的影
- • 德国、捷克草业科学学者长期放牧实验:异质草地斑块中的土壤有机碳储量和地下生物量