博文
神奇的设计-为什么基因组DNA复制通常需要RNA引导?(系列之24)
||
基因组中DNA复制最常见的情景是需要一段RNA作为“Primer”(当然个别的情况也用tRNA或蛋白质,但最常见的是一小段RNA)。对此有人只是简单地理解是因为负责催化的DNA聚合酶不具有“从头”合成的能力,而只能借助一个分子上羥基催化延伸,通过亲核取代反应形成3'-5'磷酸二脂键。其实这是造物主一个“神奇的设计”!
一、造物主选择了在细胞内贮存更多的核糖核苷酸分子而不是脱氧核糖核苷酸分子
1、中心法则信息传递中的RNA转录起始
基本的看法是细胞内核糖核苷酸的“量”差不多是脱氧核糖核苷酸分子的~100倍,本质上负责催化引物合成的“引物酶”更偏好于使用脱氧核糖核苷酸,但远高于脱氧核糖核苷酸的核糖核苷酸使得核糖核苷酸更容易在“碱基选择”中胜出。那为啥要采取这种设计?原因很简单,因为大多数细胞选择了ATP, 而不是dATP作为生命的“货币”。细胞内的“分解代谢”的“大道”是“三羧酸循环”,通过不断改变脂膜两则的质子浓度产生“电位差”,以驱动ATP的合成,并维持细胞内一个特定的能荷(ATP+ADP与ATP 、ADP和AMP总和的比值近似为0.8)。于是在DNA复制时负责在特定位点打开双链DNA和RNA催化起始位点“配上”的第一个碱基(及随后RNA复合体中负责打开双链的亚基所需要的动力,和下面的DNA解旋属于一个层级)均由ATP来“开始”。为此造物主设计了一类AAA+蛋白质(和解旋蛋白酶),对于这类分子而言,ATP的水解不用于提供能量,而是用于改变其活性中心的“构象”,使其“活起来”!这在DNA复制起点处打开双链时至关重要。
当然,遗传信息传递过程中,基因转录的情况与此不同。首先,大量的ATP(高能荷状态)不是启动基因转录的指令,而是DNA复制的指令。此时的ATP被用于帮助RNA聚合酶“从头”合成RNA。在细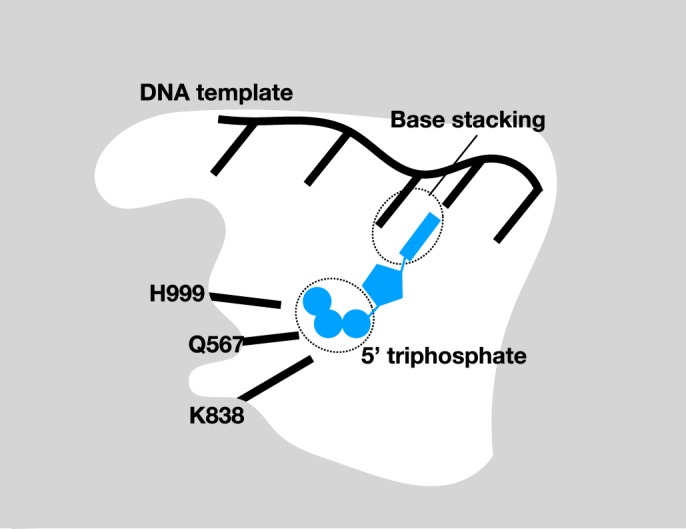
RNA聚合酶转录的RNA起始
(负责RNA转录的RNA聚合酶与负责引物RNA合成的RNA聚合活性在进化上没有关系,引物酶由拓扑异构酶进化而成)
胞内,与染色质特定位点(通常是启动子部位)结合的基础转录蛋白,如原核中的Sigma因子和真核及古细菌中的TFIID,和RNA聚合酶组成的复合体占据启动子区,然后开始转录的+1位必需用ATP与模板上的一个“T”碱基通过标准的沃森-克里克碱基配对法则配对(形成A型构象,不是B型)。
因此,用于催化转录的RNA聚合酶需要ATP结合处呈 A型构象,而不是B型构象,为此引入“碱基堆积”弱作用力,要求模板DNA上的T的3'方向碱基与A堆积,ATP中的腺嘌呤与模板上相邻的碱基形成的“碱基堆积”有助于引物酶顺利“跨越”5'A的相邻碱基,完成RNA合成的起始(这种碱基堆积的本质是因为RNA聚合酶"揪住"了核糖核苷酸ATP,并使之作为引物分子启动合成3'-5'磷酸二脂键决定,见本文附图)。
2、在DNA引物合成时RNA 分子的合成
引物酶通常由一个含有能影响DNA拓扑结构的亚基和一个RNA聚合酶或一个DNA聚合酶组成(大多数的细菌是RNA聚合酶,而真核和古细菌可以使用推进力不高的DNA聚合酶,如DNA聚合酶alpha,这为之后的DNA聚合酶之间的转换“DNA Polymerase Switching”)提供了便利。当前,包括笔者最先提出人细胞内特定染色质区的基因转录形成的RNA-DNA杂交体中RNA半分子被DNA聚合酶用于复制(已被西班牙科学家证实),及DNA形成非B型DNA二级结构后断裂引发的BIR(broken induced replication)复制 都被认为是某些与人类疾病相关的卫星DNA扩增的机制,但这属于异常情况下的复制起始。最近,发现非B结构形成后有助于促进“重启始”应该否属于类似引物酶催化的范畴。
不同物种中的引物酶(源于拓扑异构酶)均含有“Toprim”结构域,尽管引物酶中负责合成催化的“RNA聚合酶”也能催化dNTP,但此时却因为细胞核内ATP的浓度远大于dATP的浓度,所以在ATP与dATP竞争过程中ATP胜出。最终选择了ATP用于起始,这也是为什么现有细胞生命中进行基因组DNA复制时,需要合成RNA引物,而不是DNA引物的一个最合理的解释(不要把这个事件神秘化)。
目前,尚不清楚这种情形下“前导链RNA引物”的起始的分子机制与信息转录中RNA聚合酶催化起始的分子机制是否一致,但一个显而易见的区别是DNA复制需要复制出的双链DNA分子呈B型构象,而不是A型构象,这可能限制了引物RNA的长度。
二、为啥要发生DNA聚合酶和RNA聚合酶的“转换”?
一方面RNA聚合酶的3'-5'校对能力或缺如或效率低下(很多DNA聚合酶缺乏上述校对能力),因此单纯凭借基于满足新生DNA双链呈B型构象的“碱基选择”方法无法保证高的忠实性。二是DNA分子使用化学稳定性更高的脱氧核糖核苷酸,而不是稳定性差的核糖核苷酸可以保障作为遗传信息载体的DNA分子更稳定。
不能合成一次就改变了物种,也无法形成多细胞个体。基于上述原因,引物之后的DNA复制需要换酶(DNA polymerase replacement)。
三、合成冈崎片段的引物时引物酶的位置是由什么决定的?
半保留的模板选择原则使得被复制的DNA双链必须由解旋酶打开,大多数参与DNA复制的解旋酶的的构象为六元环,套住DNA单链以5'向着3'方向运动,因此解旋酶打开DNA双链的速度决定了整个DNA双链的复制快慢,而通常情况下,解旋酶背着引物酶沿着5'到3'方向的DNA上滑动(负责修复的解旋酶有些和这种负责复制的解旋酶是如蚕蠕动方向相同,有些与之相反,因此被称为5'-3'解旋酶,有的被称为3'-5'解旋酶,自当另论....
负载引物酶的解旋酶会视情况(具体原因不详)留下引物酶(是否一起驻足需要单分子模拟),于是就有了“岗崎片段”式的所谓“不连续”DNA复制,这就需要不断合成RNA引物了...
当然,DNA聚合酶只能沿着DNA模板的3'向5'方向移动,其移动速度取决于它“乘坐车”的“车速”(原核的beta夹子和真核和古细菌的PCNA滑动速度),因此DNA复制中的两条链上滑行的DNA聚合酶呈反方向,这是另外的话题了。
在这个过程中聚合酶与校对酶的转换参见我们2013年发表在The EMBO Journal 的论文。

https://wap.sciencenet.cn/blog-218980-1401649.html
上一篇:潘学峰诗选-好为难
下一篇:关于启动“呼吸爆发”或“氧化爆发”机制的假说