博文
[转载]从果蔬到乳制品,探索食品微生物对肠道健康的影响
||

俗话说病从口入,饮食对人体具有重要的影响,蔬菜和水果作为每日饮食中必不可少的成分,为人类提供了重要的营养物质,包括各种必需的维生素和矿物质。
此外,蔬菜和水果上栖息着数量惊人的微生物,高度多样化的微生物组是机会性病原体和益生菌的重要储存库。新鲜蔬菜和水果可能成为一些食源性病原微生物的重要来源。
许多研究还发现,食用新鲜蔬菜和水果会影响肠道微生物的组成,同时对人体健康和相关代谢也存在一些有益的影响,例如可以增加碳水化合物活性酶的丰度,影响短链脂肪酸和维生素的生成。
除了新鲜的蔬菜和水果外,牛奶也是人们生活中重要的饮食,牛奶富含蛋白质、维生素、矿物质、必需氨基酸等多种营养,但同时非常容易滋养微生物。不当的保存方法会促进微生物的生长和繁殖,从而对食品质量和食品安全产生负面影响。
本文从食品中的微生物角度,主要阐述了蔬果中存在的一些微生物群对肠道微生物群结构和人类健康的影响。此外还介绍了一些影响食物中微生物生长的因素,以及导致牛奶及奶制品腐败的微生物。
01
蔬果中的微生物群
人们对动物体内的微生物研究较多,有时可能忽略了植物中的微生物,你可能想象不到,一个普通的苹果就含有约1亿个细菌。
★ 蔬果中的微生物会影响人体健康
蔬果中高度多样化的微生物群是机会性病原体和益生菌的储存库。其中很少细菌是致病的,大多数是无害的,有些甚至是有益的。
随着饮食的摄入,定植于植物组织内部和外部的微生物可以在人类营养和健康中发挥作用,例如十字花科蔬菜的微生物群,研究表明它们为人类宿主提供了抵御真菌病原体和针对抗癌分子的保护。
此外,一些植物微生物与植物激素协同作用产生的代谢物,也可能与人类肠道微生物甚至整体健康相关。
▼
不同蔬果中的微生物群
研究发现蔬菜含有以放线菌门、拟杆菌门、厚壁菌门和变形菌门为主的多种细菌群落,但每种蔬菜物种的组成显著不同。
★ 肠杆菌是蔬菜微生物中的重要组成部分
这些差异通常可归因于肠杆菌科(Enterobacteriaceae)相对丰度的差异。这个革兰氏阴性菌大家族除了许多无害的共生体之外,还包括许多熟悉的机会性病原体,例如大肠杆菌(Escherichia coli)、沙门氏菌(Salmonella)、克雷伯菌(Klebsiella)。然而,肠杆菌科是蔬菜微生物组的重要组成部分。
★ 发酵的蔬菜中含有大量的乳酸菌
除了新鲜蔬菜外,发酵的蔬菜也是全世界饮食的重要组成部分,并且不同地区存在特定的传统产品,例如韩国的“泡菜”或中国的“酸菜”。
通过使用本土细菌或发酵剂进行乳酸发酵会引起细菌群落的转变。它们携带内源性乳酸菌群落,这些群落在发酵过程中变得丰富。例如,腌制橄榄含有大量乳杆菌(Lactobacillus),可能具有益生菌作用。
另一类发酵食品以奶酪和其他乳制品为代表,将在本文后面的章节讲述。
★ 不同蔬果中的微生物组成存在差异

基于宏基因组读数的分类注释表明,大部分样品中的微生物群落均以变形菌门为主,但绿橄榄除外,其中最丰富的门是厚壁菌门(占总数的69.7%)。
与其他样品相比,绿橄榄中的放线菌含量也高出10 倍(平均为4.2%对0.4%)。在属水平上,蔬菜和水果中含有不同的微生物成员(上图B所示)。
番茄中假单胞菌非常丰富
例如,假单胞菌属(Pseudomonas)是番茄中最丰富的分类群(54%),其次是拉恩氏菌属(rahnella)(22%)和泛菌属(Pantoea)(9%)。
小菠菜以泛菌属(Pantoea)(30%)为主,其次是欧文氏菌属(Erwinia)(20%)和肠杆菌属(19%)。
绿橄榄以乳杆菌(62%)为主,其次是假单胞菌(21%)、丙酸杆菌(3%)和片球菌(3%)。
黑无花果中有较高丰度的真菌
黑无花果中的细菌以紫色杆菌属(Janthinobacterium)(38%)为主,其次是泛菌属(12%)和草螺菌属(12%)。
基于从宏基因组中恢复的 16S 和 18S rRNA 基因序列的分类学图谱表明,黑无花果拥有属于真菌的最高比例的测序读数,主要分为曲霉属(30%)、未分类的散囊菌目(Eurotiales)(29%)和未分类的Trichocomaceae属(26%) 属。
注:在任何宏基因组中均未检测到古菌序列。
小结
蔬菜微生物组高度多样化,不同品种蔬菜的物种组成各不相同,并受到地理方面以及农业和食品加工的强烈影响,肠杆菌科是关键物种。
★ 蔬菜中具有潜在有益细菌和病原体
生菜在蔬菜中有着特殊的地位。它是最受欢迎的生吃蔬菜之一,并为特定微生物提供了栖息地。
研究发现,在田间种植的长叶生菜的叶际中,存在高丰度10^5–10^6菌落形成单位(cfu)g和肠杆菌科细菌比例较高的多样性。肠杆菌科分类群不仅包含潜在的有益细菌,而且还存在潜在的病原体。
在德国病原体监测系统中,在所调查的生菜样本中,1.3%发现了产维罗细胞毒素的大肠杆菌,3.8%发现了大肠杆菌。
▼
蔬菜微生物组中的条件致病菌
植物,特别是其内层和根茎,是新出现机会性病原体的重要储存库。
★ 多种病原体能够在蔬菜中定值
近年来,记录在案与食用生蔬菜有关的人类感染的数量有所增加。多种人类病原体能够在蔬菜中定殖,包括大肠杆菌等致病菌。
下图显示了细菌处理后大肠杆菌通过气孔侵入生菜叶片。
大肠杆菌在生菜叶片上的定殖
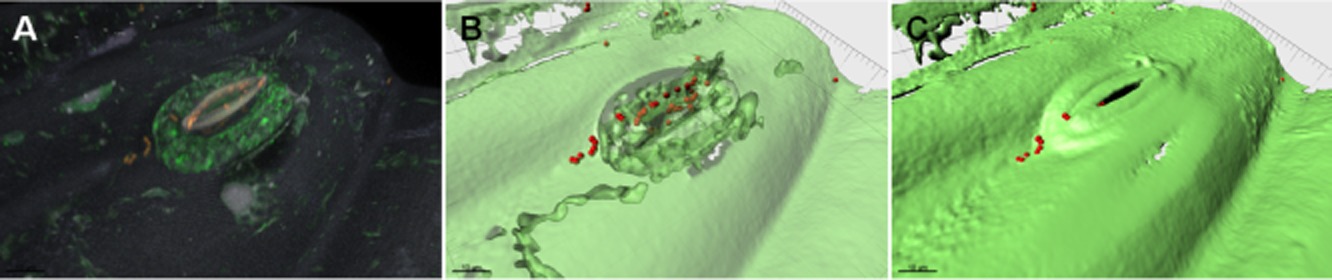
Berg G,et al.Microb Biotechnol.2014
A.共聚焦Z堆栈体积的渲染。
B和C.显示造口内细菌的A的等表面模型。
★ 植物中的一些菌属和人体存在相互作用
有许多与植物相关的菌属,包括伯克霍尔德菌属(Burkholderia)、肠杆菌属、假单胞菌属、罗尔斯通氏菌属(Ralstonia)、沙雷氏菌属、葡萄球菌属和寡养单胞菌属(Stenotrophomonas),它们与植物和人类之间相互作用。
这些属的一些成员表现出促进植物生长以及对抗植物病原体的优异拮抗特性;然而,许多菌株也会定植于人体器官和组织,从而引起疾病。一个原因是相似或通常相同的因素允许识别、粘附和入侵植物和人类宿主。
机会致病菌在芝麻菜叶层中的分类结构
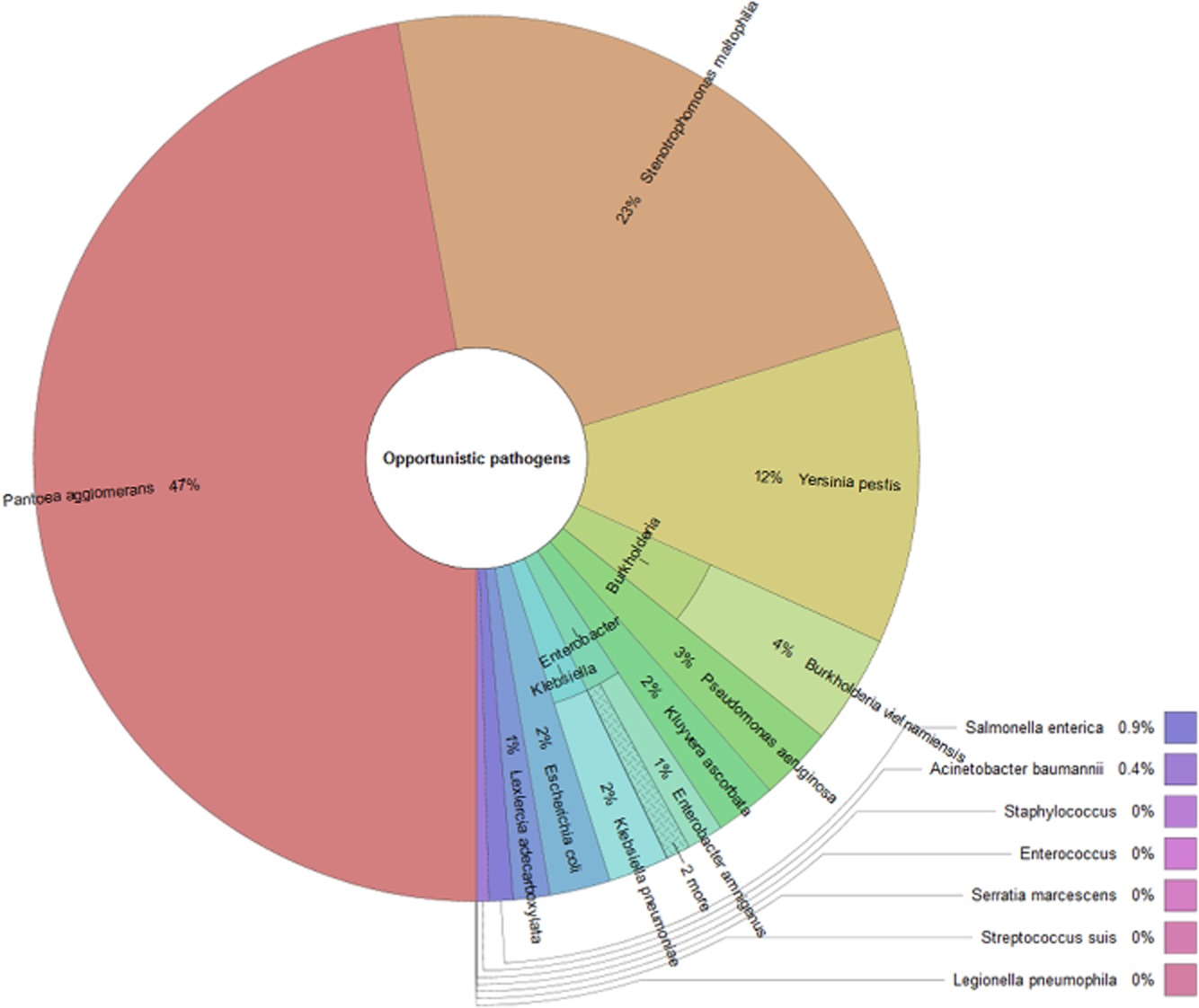
Berg G,et al.Microb Biotechnol.2014
这一群体中研究得很好的例子是革兰氏阴性,通常具有多重耐药性的铜绿假单胞菌和嗜麦芽窄食单胞菌。两者都是植物微生物群中丰富的成员,属于这些物种的菌株在基因型和表型水平上具有高度的多样性。
铜绿假单胞菌分布较广,会引起人体感染
令人惊讶的是,铜绿假单胞菌的泛基因组比人类基因组具有更大的遗传库,这解释了铜绿假单胞菌广泛的代谢能力及其在栖息地中的普遍分布。
此外,流行的植物模型拟南芥已被用于成功鉴定与毒力相关的新铜绿假单胞菌基因。
铜绿假单胞菌感染人体后,可引起病人的血液感染、肺炎、心内膜炎、尿路感染等;对植物亦是机会性感染的,感染后会流出绿色的脓汁和渗出液。
当人体自身微生物群减少时,可能被嗜麦芽窄食单胞菌感染
嗜麦芽窄食单胞菌菌株表现出相似程度的多样性。临床和环境嗜麦芽窄食单胞菌种群的多态性突变频率解释了对新生态位的适应,与植物相关的种群具有更广泛的多样性,只有突变频率高的种群(超突变者)才能适应临床环境和人类宿主。
尽管嗜麦芽窄食单胞菌菌株引起大量感染,但仅鉴定出非特异性毒力因子,例如蛋白酶和铁载体。属于该物种的菌株持续存在并表现出多重抗性;只有本土微生物群减少,病原体才有机会感染人类。
在自然栖息地中,嗜麦芽窄食单胞菌菌株定植于双子叶植物中,这些植物能够产生多种次生抗菌代谢物。
为了在这样的植物中生存,需要使用外排泵,这也是它们对临床使用的抗生素具有多重耐药性的原因。然而,研究表明,菌株水平上的基因组和表观基因组具有高度可塑性和特异性,这可能有助于强毒菌株的发展。
▼
现代工业对植物中微生物的影响
农业技术和加工对植物相关微生物群落的组成有重要影响。在过去的二十年里,更大规模的生产和更有效的新鲜蔬菜分配导致了疾病爆发数量的增加。
有机农业实践可能与传统农业实践不同,包括使用的肥料和农药的类型,这些差异有可能影响与蔬菜相关的微生物群落结构;它们通常具有较高的微生物多样性的特点。
★ 抗生素和农药的使用增加了相关耐药基因
在过去的几十年中,抗生素在畜牧业中的使用极大地促进了农场环境中抗生素耐药性的发展和丰富。特别是,粪便是耐药细菌和抗生素化合物的储存库,其应用于农业土壤被认为会显著增加抗生素耐药基因和土壤中耐药细菌种群的选择。
这些种群可以从根际侵入植物的内部,并进入人类的食物链。新鲜产品的病原体污染可能起源于收获之前或之后,一旦被污染,产品就很难消毒。
★ 动物粪便中携带的病原体可能增加了蔬菜中的病原体
食品加工也会对蔬菜微生物组的结构和食品安全产生重要影响。例如,中等干扰(例如轻微的生物或非生物影响)可以增加肠杆菌科细菌的相对丰度。
与志贺氏菌(一种以人类为主要宿主的有机体)收获前污染相关的爆发相对较少,而与沙门氏菌或产生志贺毒素的大肠杆菌相关的爆发相对较高,以动物为主要宿主的生物体,强调了家养动物和野生动物作为沙拉等蔬菜收获前污染的主要来源的作用。
既然植物中存在如此丰富的微生物组,那么其在植物生长的时候是否存在一定的作用,当人类将其作为食物摄入时又是否会对人体健康产生影响?在这一章节谷禾将具体讲述植物中微生物的相关作用。
02
植物中微生物的相关作用
▼
对植物的影响
植物微生物组对植物生长和健康起着重要作用,取决于植物种类、栽培品种和土壤类型等因素。
▷ 植物中的微生物会促进营养吸收
植物中的一些微生物可以支持营养吸收并产生多种植物激素。
例如根瘤菌和固氮菌,能够与植物共生,将大气中的氮转化为植物可利用的氮化合物。这种共生关系使植物能够吸收到更多的氮,从而促进其生长。
溶磷细菌和真菌,能够分解土壤中的有机磷化合物,将其转化为可溶性的无机磷,为植物提供营养。
一些微生物能够产生植物生长激素,如生长素和激动素,这些激素可以促进植物的生长和发育。微生物通过与植物根系接触,释放这些激素,从而影响植物根系的吸收能力。
注:激动素是一种内源的细胞分裂素,除具有促进细胞分裂的作用外,还具有延缓离体叶片和切花衰老,诱导芽分化和发育及增加气孔开度的作用。
▷ 一些植物中的病原菌会影响产量
许多病原体会对植物产生负面影响,特别是真菌、卵菌和线虫;据估计,全球三分之一以上的产量损失是由它们造成的。
病原菌会直接侵害植物的组织,破坏植物的生长和发育过程。例如,霉菌和真菌可以感染植物的叶片、茎和根部,这会削弱植物的光合作用和养分吸收能力。
此外,病原菌还可以分泌毒素,对植物产生毒害作用。这些毒素会破坏植物的细胞结构和功能,干扰植物的正常代谢过程,使植物受到氧化应激和细胞死亡的影响。
最终导致叶片枯萎、果实腐烂、凋萎和死亡等症状,从而降低产量。
▼
对人体的影响
植物微生物可能还通过刺激人体的免疫系统和增强肠道微生物组的多样性对人类健康发挥积极作用。
最近,通过与肠杆菌科细菌的显著相互作用证明了细菌多样性与特应性之间的相关性。研究发现健康人外周血单核细胞中不动杆菌(Acinetobacter)的丰度与白细胞介素10的表达呈正相关。
白细胞介素10是一种抗炎细胞因子,在维持对无害物质的免疫耐受性方面发挥着核心作用。
▷ 植物相关微生物可能成为“天然疫苗”
已知源自革兰氏阴性菌(例如肠杆菌科)的内毒素具有过敏保护和免疫调节潜力。如果植物是肠杆菌科细菌的天然储存库,那么这些细菌一定长期以来一直是我们饮食的“天然”组成部分。
考虑到全世界人们食用的蔬菜和水果量,植物相关微生物组可能可以作为免疫刺激剂或“天然疫苗”。
▷ 植物相关细菌影响肠道微生物的多样性
在人体肠道中可检测到水果和蔬菜相关细菌
有趣的是,植物和人类肠道微生物组在物种组成和功能方面存在重叠。最近的研究表明,胃并不像以前认为的那样对微生物通道构成严格的屏障。它栖息着许多物种。
最近还提供了食源性微生物(包括动物性和植物性饮食)在通过消化系统后仍能存活的证据,并且食源性菌株可能在肠道中具有代谢活性。
植物和人类相关细菌的分类及其在人类肠道中的丰度
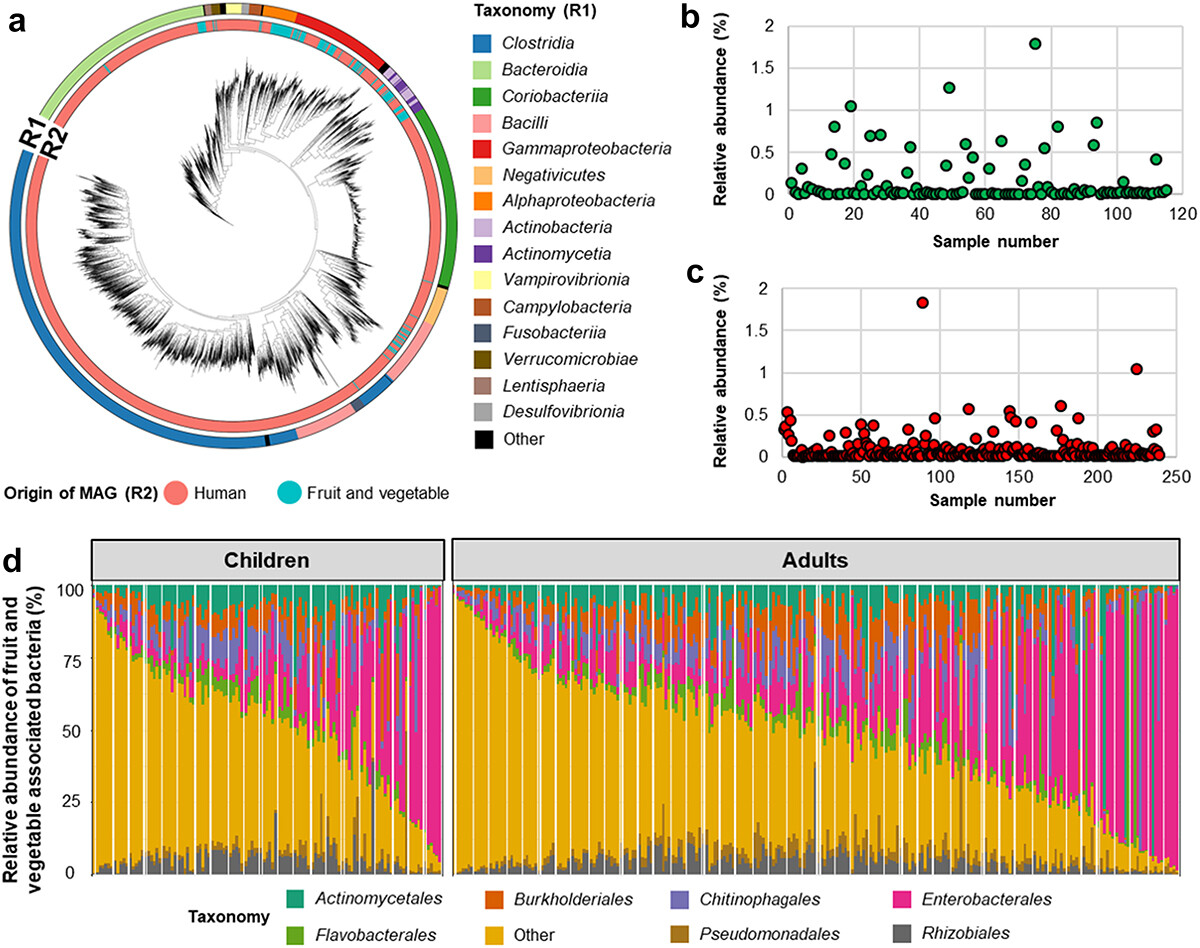
Wicaksono WA,et al.Gut Microbes.2023
(a)R1中的不同颜色表示细菌分类,R2表示MAG(宏基因组组装基因组)的起源。
水果和蔬菜相关细菌的相对丰度及其在儿童(b)和成人(c)中的丰度。条形图显示了人类肠道中与水果和蔬菜相关的细菌组成在目水平上的相对丰度。
(d)水果和蔬菜相关细菌的相对丰度是通过使用包含仅映射到水果和蔬菜相关细菌基因组读数的数据集获得的。不同的颜色代表不同的级别分类,不太丰富的细菌目包括在“其他”中。
通过基于每月粪便样本鸟枪宏基因组测序(对儿童肠道微生物组进行纵向研究,进一步尝试确定水果和蔬菜相关细菌对人类肠道整体微生物多样性的贡献。
婴儿时期喂养蔬果有助于肠道微生物的发育
有趣的是,水果和蔬菜相关细菌的多样性从1个月大到12个月大的受试者有所增加,但从此时开始下降,在24个月以上的受试者中达到最低水平。
除了母乳喂养之外,引入固体食物(即水果和蔬菜)是生命早期的事件,有助于肠道微生物组的变化和发育。
在队列中,一些儿童在前4个月内已经接触了根类蔬菜(n=269中的71名)、水果(n=269中的81名)和蔬菜。此外,制备自制婴儿食品的常见烹饪方法可能无法完全消除植物微生物群。
由于食源性微生物可以在消化道中生存,我们推测,儿童肠道宏基因组中检测到的植物相关细菌数量的增加是由于断奶早期食用的自制或生水果或蔬菜中的植物微生物群的转移和定植所致。
水果和蔬菜相关细菌平均占2.2%(最小值:0.8%,最大值:13.6%)
▷ 植物微生物随着时间的推移在体内持续存在
人类肠道中与水果和蔬菜相关的细菌群落随着宿主年龄的变化而变化,根据图中的年龄梯度,可以看到清晰的聚类。评估表明,宿主年龄影响水果和蔬菜相关的细菌群落结构(P=0.001),但只能解释5.3%的细菌变异。
有趣的是,婴儿的国籍也对水果和蔬菜相关的细菌群落结构产生了较小程度的影响(P=0.001,R2=2.2%)。这一结果可能是由于不同国家的婴儿之间的辅食喂养模式不同所致。
沿梯度富集的细菌类群主要是革兰氏阳性菌,尤其是布氏乳杆菌、乳杆菌属和乳球菌属,它们取代了早期占主导地位的泛菌属、欧文菌属和不动杆菌属。
注:与临床菌株相比,源自植物的聚集泛菌菌株同样能够定植于人类宿主。
肠道微生物的变化可能由于氧气浓度和食物种类
我们假设,由于新生儿肠道中存在氧气,来源于水果和蔬菜的兼性厌氧γ-变形菌,即泛球菌、欧文氏菌和不动杆菌,可以在断奶早期的婴儿肠道中定植。然后,由于氧气浓度降低,厌氧细菌,即布氏乳杆菌和乳杆菌属,在第8-12个月出现。
在婴儿期,肠道微生物群的生态演替是一个动态过程,然后在儿童期达到稳定阶段。随着宿主年龄的增加,人类肠道中与水果和蔬菜相关的细菌组成变得更加异质(即相对于中位数分布的差异更大)。这可能是由于通常随着婴儿年龄的增加而提供的食物种类更加多样化,也可能因个体而异,导致观察到与宿主年龄相关的异质性。
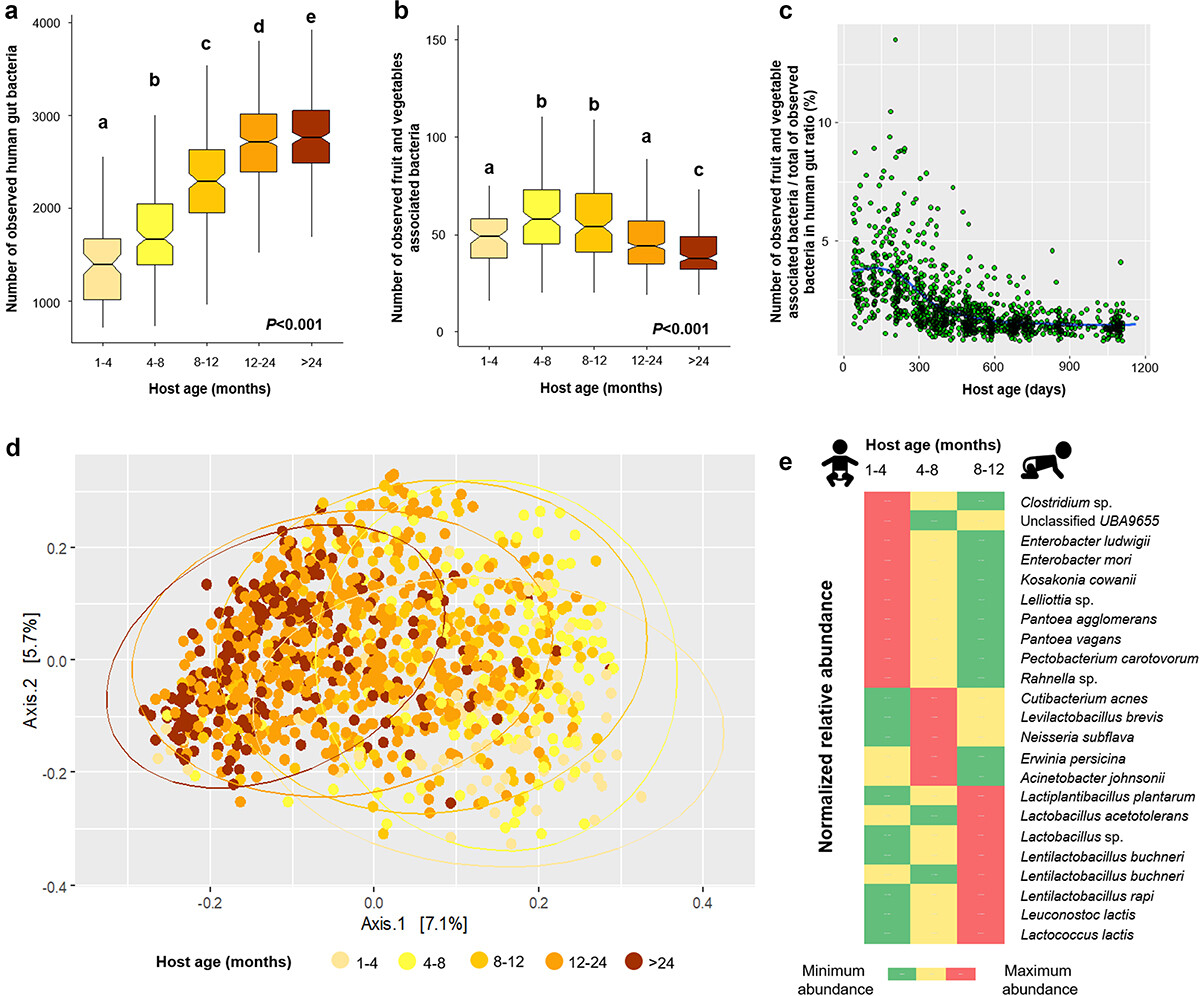
Wicaksono WA,et al.Gut Microbes.2023
箱线图包括基于观察到的肠道宏基因组观察到的人类肠道相关细菌(a)和水果和蔬菜相关细菌(b)。
箱形图上方的不同字母表示不同宿主年龄内的Kruskal-Wallis检验,然后进行成对比较(P<0.05)。散点图显示了观察到的水果和蔬菜相关以及人类肠道相关细菌宏基因组组装基因组沿年龄梯度的比率(c)。使用二维PCoA图并基于 Bray-Curtis 相异矩阵(d)显示了人类肠道中水果和蔬菜相关细菌的群落聚类。
LEfSe分析表明,水果和蔬菜相关的细菌宏基因组组装基因组在不同年龄组中富集(e)。
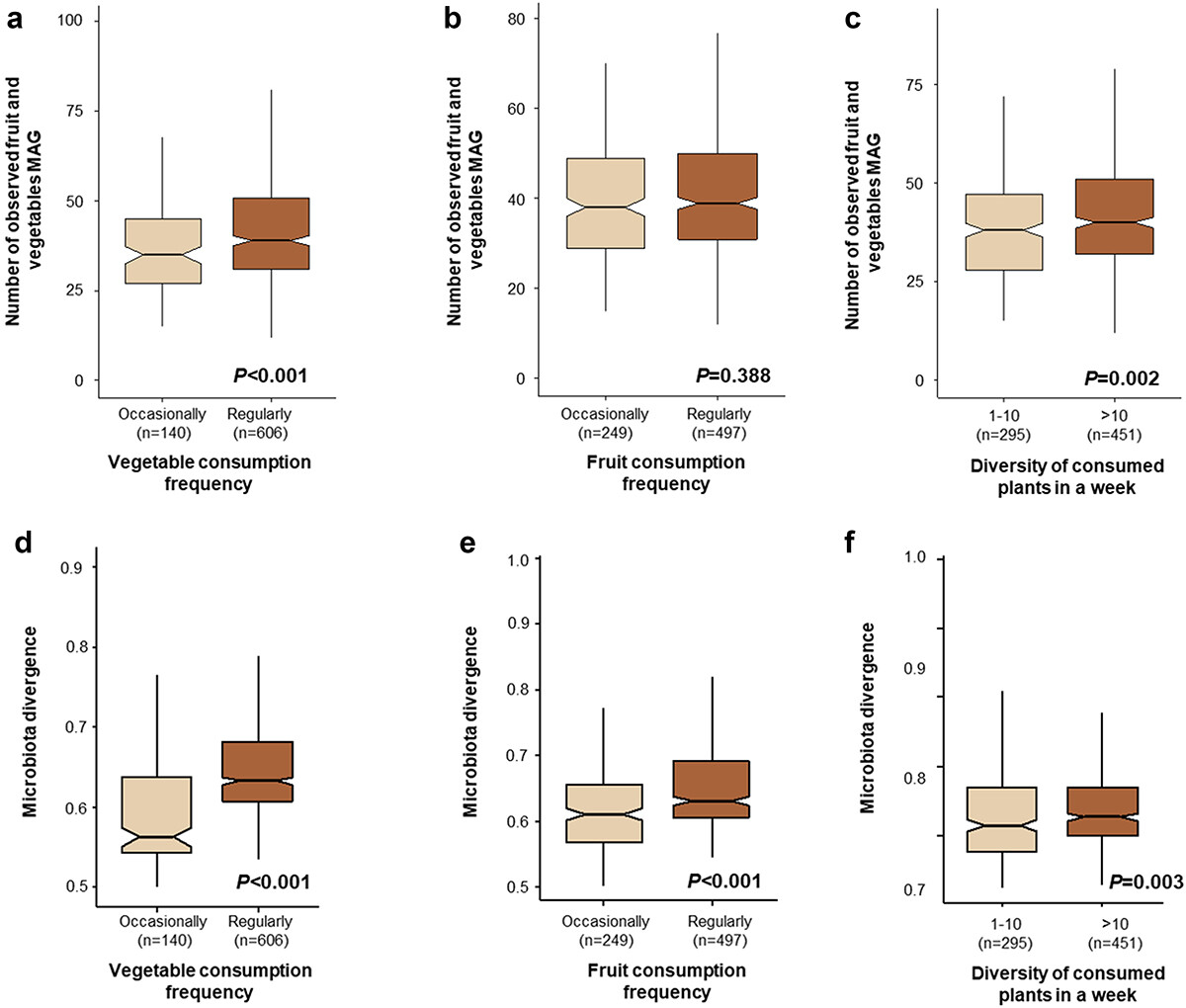
▷ 蔬果消费频率和多样性会影响人类肠道中与水果和蔬菜相关的细菌丰富度
食用水果和蔬菜的频率和多样性可能会影响人类肠道中与水果和蔬菜相关的细菌丰富度。
为了检验这一假设,研究人员使用了美国肠道项目的数据集,这是一项大型公民科学开放平台研究,收集了自我报告的饮食数据和粪便样本。
使用Kraken2,总共746个样本的读数超过500000个,这些读数被指定为细菌。保留这些样本进行进一步分析,以检查肠道微生物多样性与植物消费频率和多样性之间的关联。
分析表明,人类肠道中水果和蔬菜相关细菌的丰富度与受试者食用蔬菜的频率和多样性有关。
食用蔬菜更丰富的人群肠道微生物多样性更高
对于α多样性分析,将数据集二次采样至500000个读数后,计算了人类肠道中检测到的水果和蔬菜相关宏基因组组装基因组(MAG)的数量。
与其他组相比,经常食用蔬菜和每周食用超过10种植物的受试者中检测到的与水果和蔬菜相关的宏基因组组装基因组数量更高。然而,水果消费频率对检测到的相关宏基因组组装基因组的数量没有显著影响。
蔬菜消费频率的增加和所消费植物的多样性也增加了人类肠道中水果和蔬菜相关细菌的相对丰度。
Wicaksono WA,et al.Gut Microbes.2023
一周内蔬菜消费频率、水果消费频率和消费植物多样性对观察到的水果和蔬菜相关细菌数量以及水果和蔬菜细菌群落组成总体异质性的影响。
箱线图显示了肠道(ac)中观察到的与水果和蔬菜相关的宏基因组组装基因组。其他图显示了肠道中水果和蔬菜相关宏基因组组装基因组组成的总体异质性(df)。
与研究结果一致,经常食用蔬菜、水果和自制发酵乳制品的农村贝都因人的细菌多样性明显高于城市沙特人,城市沙特人每周只吃1-2次蔬菜和水果。
另一组研究人员也提出了类似的观点,患者因每天食用帕尔马干酪而在人类肠道中发现了与奶牛相关的细菌定植。
▼
植物中微生物群对代谢的影响
基因本体论(GO)生物学过程的预测表明,宏基因组读数的很大一部分与代谢功能有关,如核酸代谢、蛋白质代谢和碳水化合物代谢。
★ 植物中不同的微生物组成导致代谢存在差异
对四个样本中注释基因的比较表明,并非所有生物代谢过程都是均匀分布的。在新鲜菠菜中,与其他样品相比,我们发现分配给果胶分解代谢、甘油醚代谢、海藻糖生物合成、L-苏氨酸分解代谢为甘氨酸和L-丝氨酸生物合成过程等过程的高频读取。
在无花果中,我们发现木葡聚糖代谢和聚羟基丁酸酯生物合成更加频繁。与其他样品相比,番茄中的乙醛酸分解代谢更为频繁。
此外还确定了与人类肠道微生物组稳态相关的代谢途径的存在,包括谷氨酸、萜类化合物、核黄素(VB2)、硫胺素(VB1)、叶酸、谷氨酰胺和生物素的生物合成。
除绿橄榄外,所有样品中都存在类胡萝卜素生物合成过程,而对氧化还原状态、乳糖代谢过程以及脂磷壁酸、磷壁酸和异戊烯基二磷酸生物合成过程的响应仅在绿橄榄中被发现。
总体而言,这些数据揭示了食用植物微生物组具有多样化的基因组库,且不同样本之间存在差异。
▷增加碳水化合物活性酶的丰度
为了进一步了解这些植物微生物组的功能,我们研究了碳水化合物活性酶 (CAZymes) 的丰度,这是一组参与植物细胞壁分解的酶,有可能进一步转化为人体肠道中的有益代谢物。
碳水化合物活性酶根据蛋白质序列和结构相似性进行分类,包括糖基转移酶(GTs)、糖苷水解酶(GHs)、碳水化合物酯酶(CE)、多糖裂解酶(PL)、辅助活性酶(AA)和碳水化合物结合模块(CBM)。
本研究中所表征的植物相关微生物群中的假定碳水化合物活性酶是使用来自宏基因组和碳水化合物活性酶数据库的预测开放阅读框进行鉴定的。
糖基转移酶和糖苷水解酶是宏基因组中预测最丰富的碳水化合物活性酶(约占总数的30%至57%)。样品中鉴定出较少百分比的辅助活性酶(约1-6%)、碳水化合物结合模块(2-5%)、碳水化合物酯酶(3-9%)和多糖裂解酶(1-4%)。
具体而言,碳水化合物活性酶数据库中的133个糖苷水解酶家族中有79个在样本中被检测到,其中 GH13、GH23和GH1是最常见的家族。GH13是人类肠道微生物组中最常见的家族之一,其特点是分解淀粉。
部分糖苷水解酶与人类肠道的有益作用相关
研究还观察到糖苷水解酶明显分布在特定样本中,并且与人类肠道的有益作用相关。
例如,番茄酶与木聚糖酶一起是GH10的成员。番茄红素酶促进抗真菌番茄红素降解为非致癌底物。
黑无花果中真菌的存在可能与GH10酶的存在有关,GH10酶使真菌毒素呈惰性。
GH1包含参与纤维素降解的β-葡萄糖苷酶,纤维素是叶类蔬菜中最丰富的纤维之一。肠道微生物利用GH1酶将聚糖水解成葡萄糖和糖酵解前体葡萄糖-6-磷酸。
含有海藻糖磷酸化酶的GH65在绿橄榄样品中含量最高。海藻糖是一种二糖,可以防止植物干燥,存在于橄榄叶中。海藻糖酶也存在于脊椎动物的小肠中,肠球菌等细菌已被证明可以代谢海藻糖。
GH28家族含有与果胶降解相关的酶。果胶被认为是一种益生元,番茄中存在的欧文氏菌属(Erwinia)和假单胞菌属(Pseudomonas)是常见的果胶分解酶生产者。
总之,这些数据进一步支持了这样一种观点,即植物中存在的微生物在作为饮食的一部分食用时可以为人类提供代谢益处,因为它们含有大量碳水化合物活性酶,对人类健康和植物细胞壁中存在的底物起着至关重要的作用。
▷ 影响短链脂肪酸产生的代谢途径
与人类健康相关之一的微生物代谢物是短链脂肪酸。短链脂肪酸源自复杂碳水化合物的微生物发酵,既可作为重要的调节信号分子,又可作为宿主的额外燃料来源。
所分析的四个样品中最普遍的短链脂肪酸途径是乳酸氧化和L-赖氨酸分解代谢过程为乙酸。在新鲜菠菜和绿橄榄的宏基因组中也发现了乳酸代谢过程。最不常见的途径是丁酸代谢过程,因为宏基因组包含的丁酸激酶读数最少。
蔬果中的微生物组有助于产生乙酸和丙酸
有趣的是,研究中具有最广泛短链脂肪酸代谢潜力的样品是腌制的绿橄榄,这表明腌制过程可能会选择具有广泛短链脂肪酸生产能力的微生物。样品中最具代表性的酶是与乙酸(乙酸激酶)和乳酸(D-乳酸脱氢酶)生产相关的酶。
这些数据表明,水果和蔬菜的微生物组含有产生乙酸和丙酸(与人类健康相关的微生物代谢物家族)所需的基因和途径,但缺少丁酸生物合成,因为它主要与样品中未见的严格厌氧菌有关。
▷ 与维生素合成相关
众所周知,微生物可以合成维生素,这对于各自宿主的多种代谢反应至关重要。
本研究的宏基因组中参与维生素生物合成和摄取转运蛋白酶的鉴定是基于先前在人类粪便宏基因组和益生菌菌株克劳氏芽孢杆菌(Bacillus clausii)中鉴定的KEGG同源物(KOs)注释基因。
植物微生物中存在编码维生素合成的基因
研究结果表明,植物相关宏基因组中存在编码酶的基因,这些酶是钴胺素(维生素B12)、生物素(维生素H)和泛酸(维生素B5)生物合成途径的一部分,叶酸(维生素B9)、甲基萘醌(维生素K2)和烟酸(维生素B3)生物合成的基因簇较少。
此外,在宏基因组中检测到了很高比例的与钴胺素代谢相关的基因,并且在早期番茄和绿橄榄中发现了略高丰度的钴胺素代谢基因。
钴胺素(维生素B12)是人类必需的辅助因子,由一些细菌和古细菌合成,参与介导微生物-微生物相互作用和宿主-微生物作用。钴胺素缺乏会导致贫血、神经系统疾病等。
03
影响食物中微生物生长的因素
植物和动物来源的食物都在微生物生长中发挥着重要作用。微生物在食品中生长或繁殖的能力取决于食品环境。
食品的内在因素、外在因素、隐性因素包括各种食品加工方法分别对微生物的生长产生作用。这些因素决定了食物中微生物的生长以及它们用来产生能量和代谢产物的特定途径。

Part 01
内在因素
1
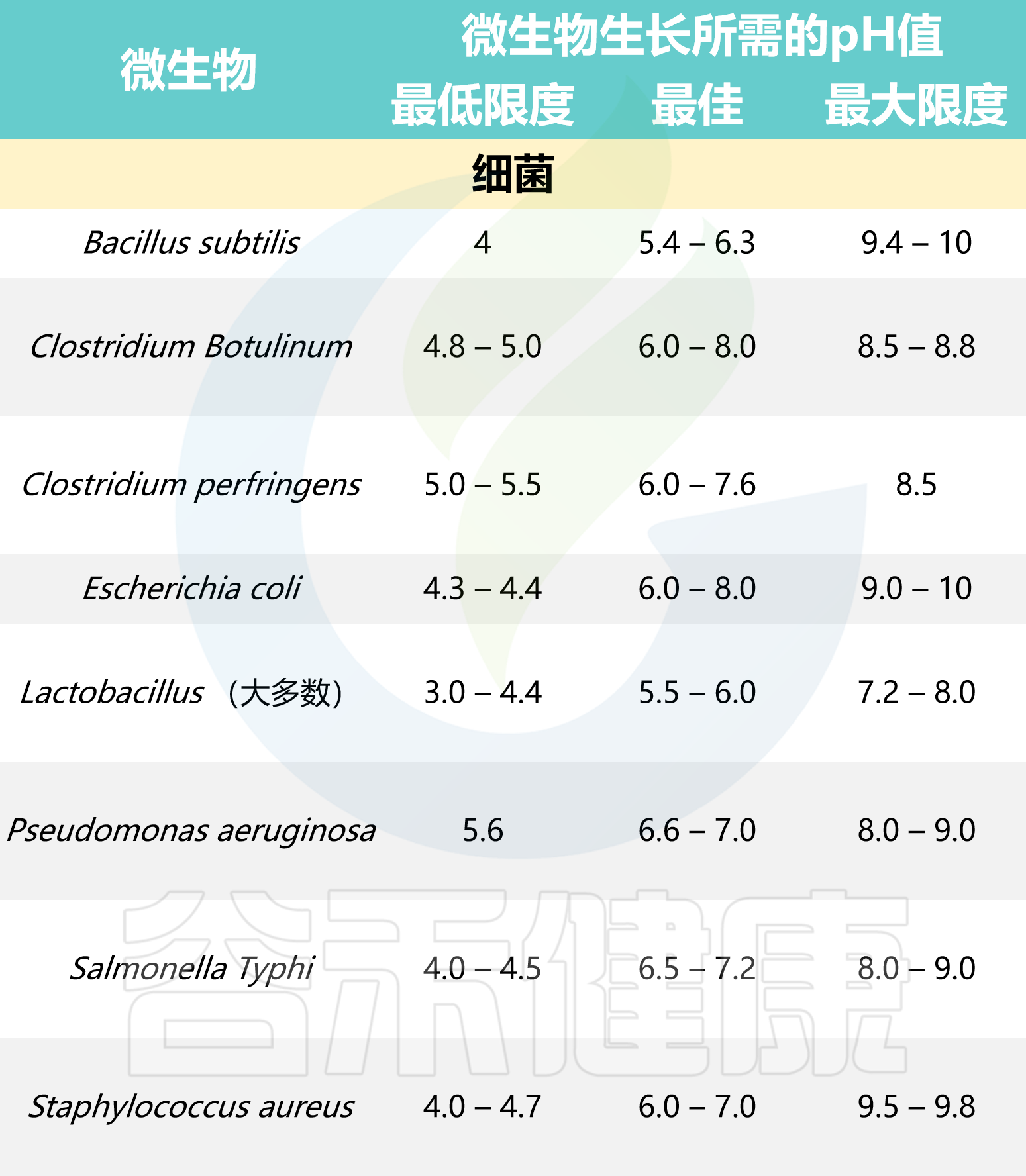
酸碱度
一般来说,与细菌相比,霉菌和酵母菌可以在较低的pH值下生长,并且革兰氏阴性菌比革兰氏阳性菌对低pH值更敏感。
霉菌生长的pH范围为1.5至9.0;对于酵母生长的pH范围为2.0至8.5;革兰氏阳性菌生长的pH范围为4.0至8.5;革兰氏阴性菌生长的pH范围为4.5至9.0。
根据 pH 范围,微生物可分为:
中性菌在pH值5至8范围内生长最佳;
嗜酸菌在pH值低于5.5时生长最佳;
嗜碱菌在pH值高于8.5时生长最佳。
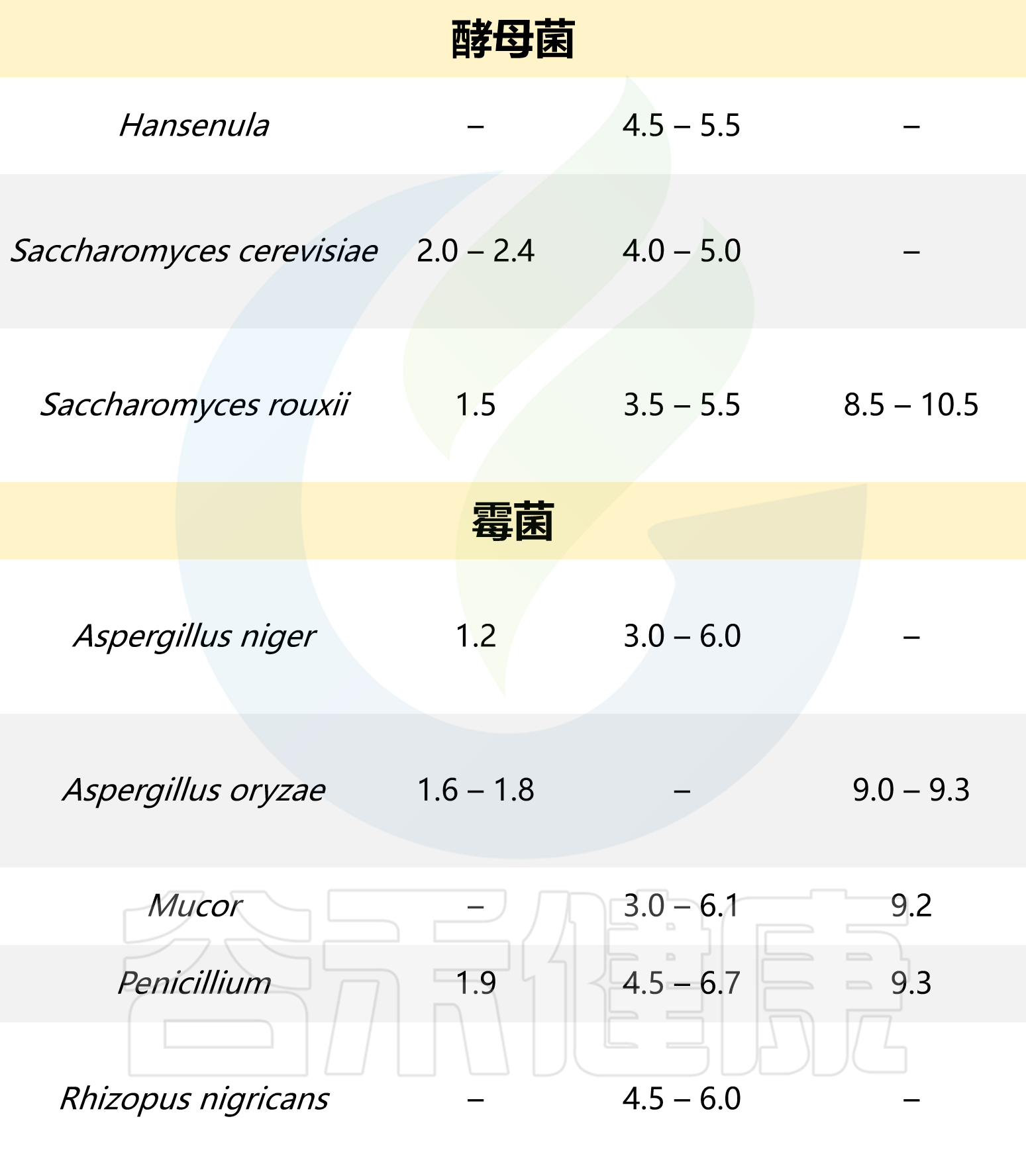
以下是一些常见微生物生长所需的pH值:
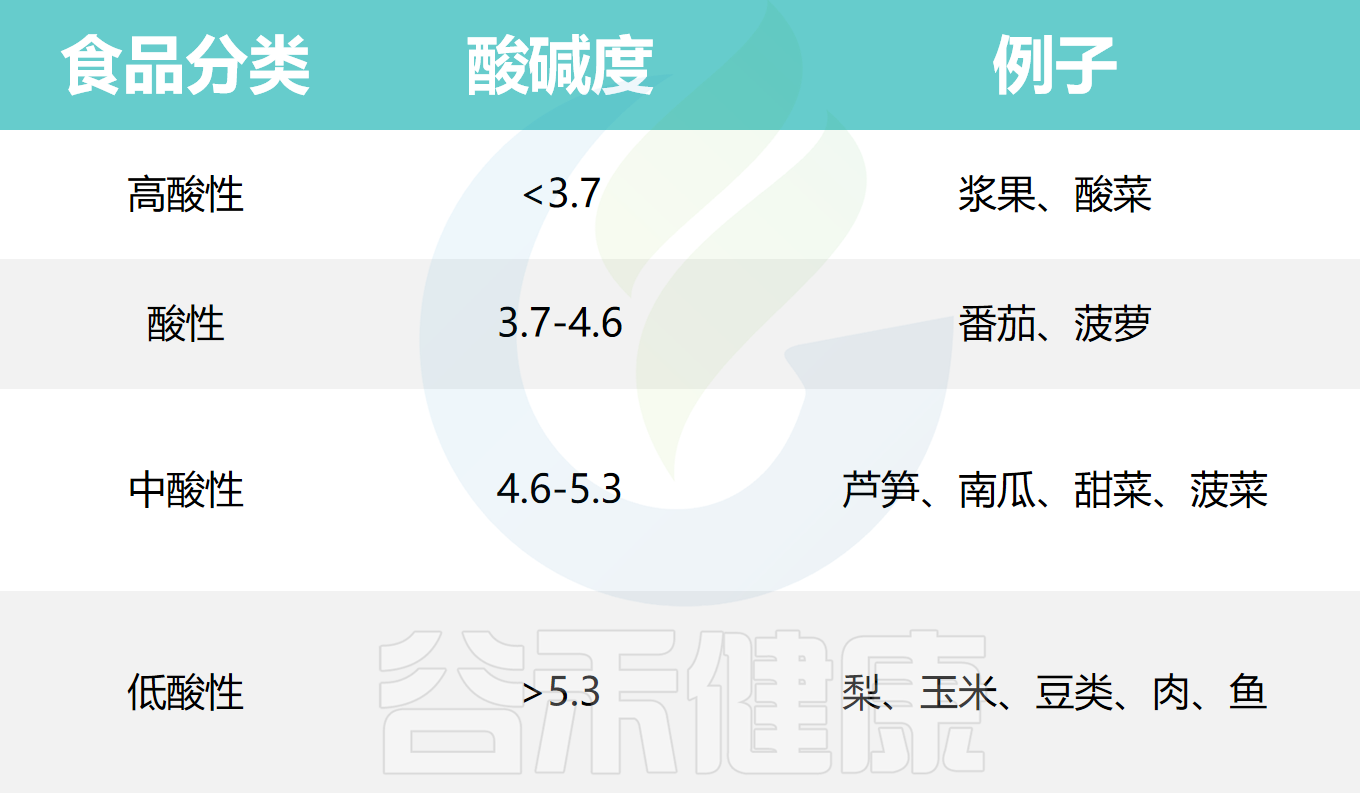
根据pH值,食物一般可分为以下几类:
2
水分活度
水分活度(aw)是可用于生物功能的水量,可因渗透效应而减少。微生物需要可用形式的水才能在食物中生长。
食物的水分活度可以用相同温度下食物的水蒸汽压与纯水的水蒸汽压力的比值来表示,0<它的范围是<1。
与真菌相比,细菌的生长需要更高的水分活度。细菌不会在低于0.91的水平生长,霉菌可以在低至0.80的水平生长。革兰氏阴性菌比革兰氏阳性菌对低水分活度更敏感。
3
氧化还原电位
物质的氧化还原或氧化还原电位被定义为原子或分子之间电子转移的测量值。氧化还原电位通常写为Eh,并以毫伏(mV)为单位进行测量。
不同类群微生物生长所需的Eh范围如下:
需氧菌在+500至+300mV下生长最佳,如霉菌、酵母、芽孢杆菌、假单胞菌、卡他莫拉菌和微球菌;
兼性厌氧菌在+300至+100mV下生长最佳,例如乳酸菌和肠杆菌科细菌;
厌氧菌在+100至–250mV或更低的电压下生长最佳,例如梭菌属。
4
营养成分
微生物的生长和代谢功能需要蛋白质、碳水化合物、维生素、脂质、水、以及硫、磷、氮等矿物质。
人们的食物是微生物生长的最佳营养来源,但食品中常见的微生物对营养的需求差异很大,能够大量利用现有营养的微生物通常在食品中占主导地位。
首先利用简单的碳水化合物和氨基酸,然后是这些营养素的更复杂形式。革兰氏阳性菌的营养需求高于酵母菌,其次是革兰氏阴性菌,霉菌的营养需求最低。
因此许多食物中都会出现霉菌
5
抗菌成分
部分食品具有天然的抗菌成分,这些成分可以抑制微生物的生长和繁殖。因此其中一些可以用作天然的保存剂。
下面罗列了一些常见食物中的抗菌成分:


6
生物结构
有些食物具有防止微生物进入的生物结构,天然覆盖物可防止损坏并减少微生物腐败的机会。
例如水果的外壳、坚果壳、蛋壳、肉类具有筋膜和皮肤等结构,可以防止食源性病原体和腐败微生物的进入。
Part 02
外部因素
受外部条件控制的因素称为外在因素,这些因素包括温度,相对湿度等。
1
储存温度
酶促反应和微生物生长受到环境温度的影响。例如酵母菌和霉菌的生长温度范围很广,为10–35°C。
根据温度,细菌通常可分为以下几类:
-嗜冷菌可以在0-20°C的温度范围内生长,例如假单胞菌属(Pseudomonas)和肠球菌属(Enterococcus)。
-嗜温菌是可以在25°C-40°C之间生长的微生物,最适生长温度接近37°C。 这些包括沙门氏菌、葡萄球菌、梭菌、志贺氏菌。
-嗜热菌是在45°C以上高温下生长的微生物,最适生长温度在50°C-70°C之间。其中包括芽孢杆菌属、梭菌属和地热芽孢杆菌(Geobacillus)。
注:病原菌一般均为嗜温菌,最适温度为人体的体温,即37℃,故实验室一般采用37℃培养细菌。
2
相对湿度
相对湿度是大气或食品环境中的水分含量。相对湿度会影响食品的水分活度水平,从而影响微生物的生长。
低湿环境(相对湿度低于30%):低湿环境会导致细菌细胞失水,从而抑制其生长和繁殖。细菌在干燥环境中往往处于休眠状态,无法进行正常的代谢活动。因此,低湿环境对于细菌的生长是不利的。
中等湿度(相对湿度在30%至70%之间):中等湿度是细菌生长的最适宜条件之一。在适度湿润的环境中,细菌可以获得足够的水分来维持其细胞活动,并且可以更好地利用营养物质进行生长和繁殖。因此,中等湿度下的细菌生长速度较快。
储存在较高湿度环境中的干燥谷物会吸收水分并发生霉变。
3
气体浓度
气体浓度可以对微生物的生长产生影响。不同的微生物对气体浓度有不同的适应能力和偏好,因此气体浓度的变化可能会对微生物的生长速率和代谢活性产生影响。
以下是一些常见气体对微生物生长的影响:
-氧气:氧气是许多微生物生长所必需的,被称为好氧微生物。它们需要氧气进行呼吸作用,以产生能量和完成生物化学反应。然而,有些微生物是厌氧微生物,它们在缺氧或无氧条件下生长。氧气浓度过高会抑制厌氧菌的生长甚至导致其死亡。
-氮气:氮气在大气中的浓度很高,但大多数微生物无法直接利用氮气。一些微生物具有固氮能力,能够将氮气转化为可利用的氨或亚硝酸盐,从而为其他微生物提供可用的氮源。
-甲烷:甲烷是一种重要的温室气体,也是一些微生物的产物。产甲烷菌(Methanogenus)是一类厌氧微生物,它们能够利用无机或有机化合物产生甲烷。这类微生物对甲烷浓度的变化非常敏感。
Part 03
隐性因素
第三个因素被称为隐性因素。该因素包含了生物体本身的特性以及一些食品加工手段对微生物生长的影响。
•一种微生物对其他微生物可能存在抑制作用
微生物可以抑制或刺激另一种微生物的生长。一些微生物可能会产生对其他生物体具有抑制性或致命性的物质,例如抗生素、细菌素、过氧化氢和有机酸。
•食品加工
在食品加工过程中,微生物会受到各种物理或化学压力。这些加工因素包括加热、冷冻、干燥、减少气压、辐射和各种化学物质:
加热通过破坏细胞质膜、改变代谢和酶活性来帮助降低食物中的微生物水平。
冷冻通过降低pH值和增加水分活度的抑制作用来减少微生物的生长。
干燥会减少微生物的生长,因为它会导致代谢损伤,从而损害细胞的增殖。
04
牛奶及奶制品中的微生物腐败
牛奶被称为均衡食品和高营养食品,富含蛋白质、脂肪、碳水化合物、维生素、矿物质、必需氨基酸等。是许多人日常生活中常摄入的食品之一。
但牛奶的高水分活度、适中的pH值(6.4–6.6)和高营养含量,也成为许多微生物(即细菌、病毒、真菌和原生动物)的极佳生长培养基。
★ 牛奶及奶制品非常容易受微生物的影响
牛奶及其制品,如酸奶、黄油、奶油和奶酪都很容易受到微生物的影响而腐败。
生奶和巴氏奶中都含有多种微生物,它们都是冷藏保存的,但保质期有限。
在巴氏灭菌前的冷藏(奶牛场和加工厂)期间,只有耐冷菌可以在冷藏奶中生长,例如假单胞菌(Pseudomonas)、黄杆菌(Flavobacterium)、产碱杆菌属(Alcaligenes)、单核细胞增生李斯特氏菌、小肠结肠炎耶尔森氏菌、某些大肠菌群和芽孢杆菌。
其中一部分是致病菌,会引起食物中毒,使人出现呕吐、腹泻、头晕等症状,严重时甚至可导致死亡,建议大家避免直接食用生奶。
在巴氏灭菌中存活下来的是一些耐热微生物,如微球菌、一些肠球菌、链球菌、一些乳杆菌以及梭状芽胞杆菌。霉菌和酵母菌通常在巴氏灭菌过程中被消除。
在较小程度上,原生动物病原体如隐孢子虫和贾第鞭毛虫也已被发现污染牛奶。
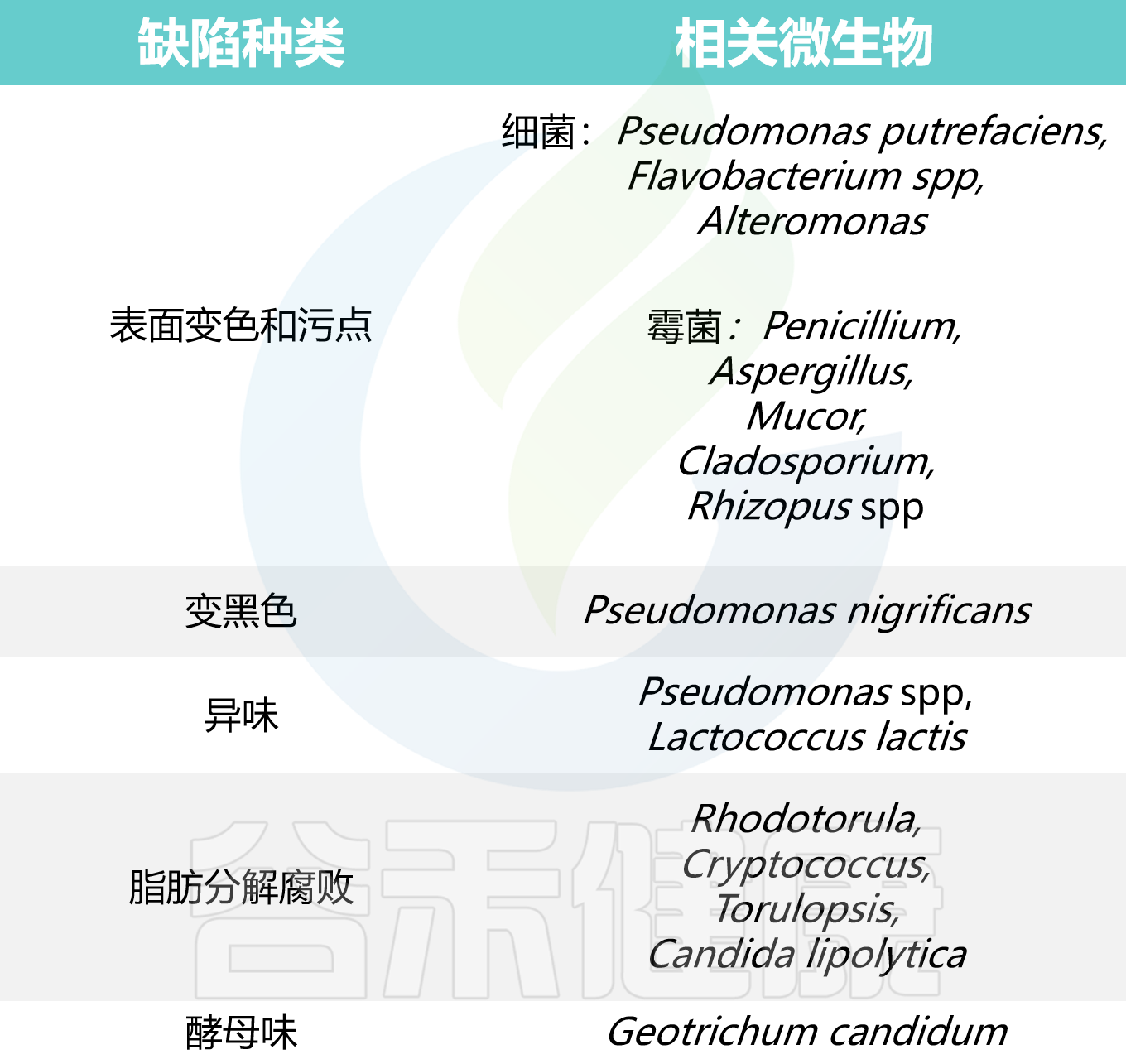
★ 微生物引起的牛奶变化
一些微生物可能引起牛奶出现以下变化:
-出现异味
-变色
-产生气体
-脂肪分解并发生酸败
-蛋白水解
-在低酸度下凝结
谷禾在下表列出了一些微生物对应的牛奶变化:

▼
乳制品的腐败变质与微生物密切相关
人们可能觉得乳制品经过加工后,不太容易变质,但事实上,在常温下许多乳制品还是会被相关微生物污染并引起腐败。
乳制品的微生物量主要取决于:
•使用的奶和奶制品的类型(生奶、炼乳、奶粉、奶油、黄油等);
•用于增强其功效的产品,如明胶、水果、糖、巧克力、着色剂等;
•各类设备的卫生水平、巴氏灭菌的效率、生产和包装过程中的卫生水平。
接下来为大家简单介绍一下引起乳制品变质的相关微生物。
•奶油变质
奶油是由均匀化前沉积在牛奶顶部的乳脂层制成的。
冷却牛奶用于生产奶油,因此嗜冷菌是腐败的主要原因,包括假单胞菌属、产碱杆菌属、不动杆菌属、气单胞菌属和无色杆菌属。
在室温下,引起奶油腐败的微生物是棒状杆菌(Corynebacterium)、芽孢杆菌(Bacillus)、微球菌(Micrococcus)、乳杆菌(Lactobacillus)和葡萄球菌(Staphylococcus)。
奶油对大肠杆菌、鼠伤寒沙门氏菌和单核细胞增生李斯特氏菌等病原微生物高度敏感。

•黄油变质
黄油是通过牛奶分离和随后的奶油搅拌制成的奶制品。奶油是黄油的原材料,因此黄油中发现的微生物主要来源是奶油。
引起黄油腐败的主要微生物是霉菌,例如Thamnidium、Cladosporium和Aspergillus。
此外,引起黄油变质的病原微生物有李斯特菌、布鲁氏菌、分枝杆菌、弯曲杆菌、小肠结肠炎耶尔森氏菌、鼠伤寒沙门氏菌等。
•奶酪变质
奶酪是一种发酵乳制品,是通过使用凝乳酶凝固牛奶中的酪蛋白而制成的。奶酪的成熟是各种微生物的蛋白水解和脂肪分解活性而实现的。
新鲜的软奶酪由于pH值较高、水分含量较高且盐度较低,因此很容易变质;硬质和半硬质成熟奶酪的水分含量较低,与细菌相比,更容易受到真菌的侵害。
细菌引起的奶酪变质
细菌性奶酪变质是由梭菌属(尤其是C.pasteurianum,C.butyricum,C.sporogenes和C.tyrobutyricum)、多粘芽孢杆菌(Bacillus polymyxa)、黄杆菌、假单胞菌、产碱杆菌和无色杆菌引起的。
真菌引起的奶酪变质
酵母菌也是引起奶酪腐败微生物之一,例如念珠菌属、汉斯德巴氏酵母菌、白地霉和毕赤酵母。由霉菌引起的腐败主要是青霉菌和枝孢菌。
奶酪中发现的主要致病菌是李斯特菌、沙门氏菌和大肠杆菌。

•酸奶变质
酸奶是牛奶经过巴氏杀菌后再添加有益菌(主要是嗜热链球菌和保加利亚乳杆菌)发酵后制成的。
霉菌和酵母菌是酸奶中的主要污染物
导致酸奶腐败的常见霉菌是曲霉属、青霉属、根霉属、镰刀菌属和木霉属。
导致酸奶腐败的常见酵母菌是念珠菌属、汉斯德巴氏酵母菌、克鲁维酵母菌、球拟酵母属。
霉菌和酵母使酸度下降,导致细菌蛋白水解和腐败,产生气体、异味和变色等。
据报道,酸奶中常见的食源性病原体有大肠杆菌、金黄色葡萄球菌、李斯特菌和小肠结肠炎耶尔森菌。
结语
食物中的微生物对于食品的口感、可消化性和安全性都起着重要作用,伴随饮食摄入的微生物可以调节肠道微生物结构甚至影响人体健康。
例如新鲜水果和蔬菜中的微生物具有大量的分类学和功能多样性,可以增加人体肠道微生物的多样性,调节碳水化合物、短链脂肪酸及维生素的相关代谢。
与此同时,蔬果微生物组是机会性病原体的储存库。在免疫功能低下的个体中,机会性病原体可引起严重感染。这些感染包括肺炎、血液感染、尿路感染、手术部位感染和腹泻等。
在免疫能力强的宿主中,植物相关微生物可能成为“天然疫苗”,刺激免疫系统并增强微生物多样性以维持我们的健康。
生活中对于食品的保存也至关重要,不当的保存方法会导致有害微生物大量繁殖,影响食物的风味及安全,进而影响人体健康。
了解食品微生物的组成和功能及其相互作用对于人体微生物多样性和系统健康非常重要。微生物检测、多组学整合可以提供技术解决方案,而益生菌、益生元和合生元可以提供改善方案。
主要参考文献
Wicaksono WA, Cernava T, Wassermann B, Abdelfattah A, Soto-Giron MJ, Toledo GV, Virtanen SM, Knip M, Hyöty H, Berg G. The edible plant microbiome: evidence for the occurrence of fruit and vegetable bacteria in the human gut. Gut Microbes. 2023 Dec;15(2):2258565.
Soto-Giron MJ, Kim JN, Schott E, Tahmin C, Ishoey T, Mincer TJ, DeWalt J, Toledo G. The Edible Plant Microbiome represents a diverse genetic reservoir with functional potential in the human host. Sci Rep. 2021 Dec 15;11(1):24017.
Berg G, Erlacher A, Smalla K, Krause R. Vegetable microbiomes: is there a connection among opportunistic infections, human health and our 'gut feeling'? Microb Biotechnol. 2014 Nov;7(6):487-95.
Abdelfattah A, Tack AJM, Lobato C, Wassermann B, Berg G. From seed to seed: the role of microbial inheritance in the assembly of the plant microbiome. Trends Microbiol. 2023 Apr;31(4):346-355.
Abdelfattah A, Freilich S, Bartuv R, Zhimo VY, Kumar A, Biasi A, Salim S, Feygenberg O, Burchard E, Dardick C, Liu J, Khan A, Ellouze W, Ali S, Spadaro D, Torres R, Teixido N, Ozkaya O, Buehlmann A, Vero S, Mondino P, Berg G, Wisniewski M, Droby S. Global analysis of the apple fruit microbiome: are all apples the same? Environ Microbiol. 2021 Oct;23(10):6038-6055.
Mendes R, Garbeva P, Raaijmakers JM. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol Rev. 2013 Sep;37(5):634-63.
Li J, Wang Z, Karim MR, Zhang L. Detection of human intestinal protozoan parasites in vegetables and fruits: a review. Parasit Vectors. 2020 Jul 29;13(1):380.
本文转自:谷禾健康
https://wap.sciencenet.cn/blog-2040048-1407214.html
上一篇:[转载]肠道菌群:开启癌症治疗协同个性化调节策略新篇章
下一篇:[转载]为什么药物对人效果不一?探索药物-微生物群相互作用对效果的影响