ВЉЮФ
ДѓздШЛШчКЮдкЕиЧђЩЯЁАЕёзСЁБГіЪ§АйЭђИіЮяжжЃП
 ОЋбЁ
ОЋбЁ
||
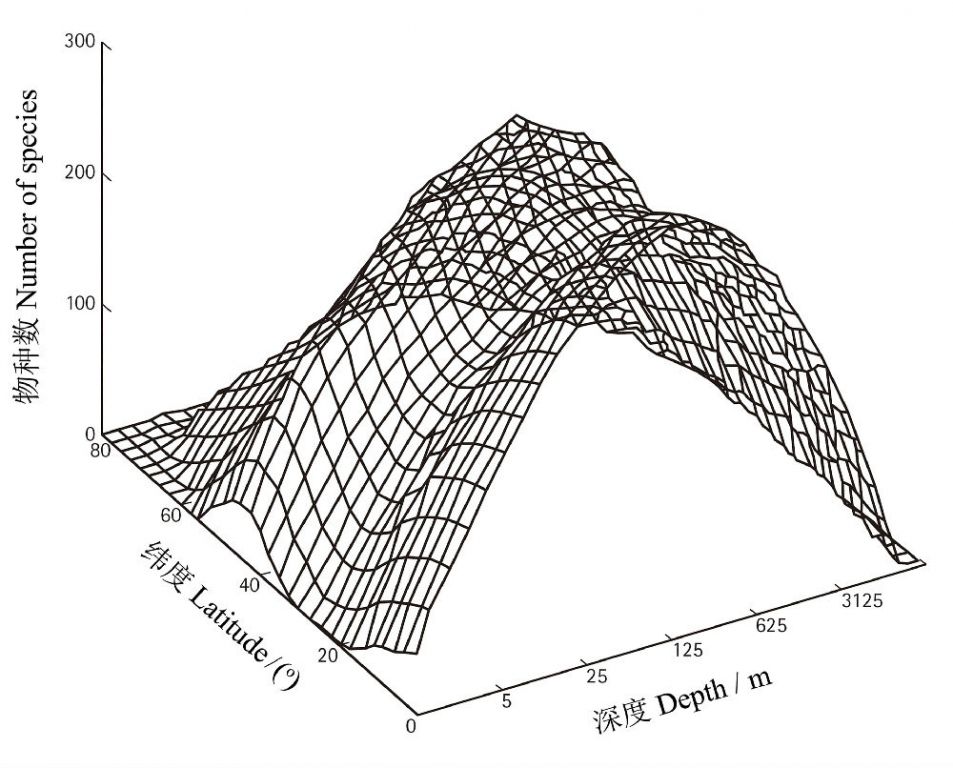
ЮяжжЪЧЩњУќЩњДцгыЗБбмЕФзюЛљБОЕЅдЊЃЌДгвЛИізюживЊЕФВрУцЃЈРњЪЗадЕиЁЂЯжЪЕадЕиЃЉядПЬЛђУшЛцзХЕиЧђЩЯЩњУќЯЕЭГЕФЖЏЬЌадЕФЖрзЫгыЖрВЪЁЃЮяжжЪЧШКТфКЭЩњЬЌЯЕЭГЕФНсЙЙЕЅдЊЃЌвВЪЧвХДЋгыНјЛЏЕФЛљБОЕЅдЊЁЃвдЮяжжЮЊЕЅЮЛЕФЩњЮябЇЁЂЩњЬЌбЇжЊЪЖвВЪЧШЯЪЖЩњЬЌЯЕЭГЙІФмЁЂЙмРэЁЂБЃЛЄгыаоИДздШЛЩњЬЌЯЕЭГВЛПЩЛђШБЕФЛљДЁ......
ВЛЩйШЫШЯЮЊНјЛЏЃЈЛђепЫЕЮяжжЗжЛЏЃЉЪЧЫцЛњЕФЁЃЕЋЪЧЃЌЪТЪЕЩЯНјЛЏМШЪЧХМШЛвВЪЧБиШЛЕФВњЮяЁЃБиШЛадНЈСЂдкетбљвЛжжЪТЪЕЛљДЁжЎЩЯЃЌМДЮяжжЪЧдкТўГЄЕФЕижЪФъДњЃЌЩњУќЖдЦјКђЛЗОГЕФЪЪгІвдМАЩњЮяМфЕФЯрЛЅзїгУЕШЪЪгІНјЛЏЖјаЮГЩЕФЁЃДѓСПЕФЪТЪЕвбОжЄУїЃЌЮяжждкИїжжВЛЭЌЕиРэЧјгђОпгаВЛЭЌЕФЗжВМЬиеїгыЖрбљЖШЃЌМДДцдкЫљЮНЮяжжЕФЕиРэИёОжЁЃашвЊжИГіЕФЪЧЃЌЮяжжЕФЕиРэИёОжВЂВЛЪЧвЛГЩВЛБфЕФЃЌЫќЪмЕНвЛЯЕСаЛЗОГвђЫиЬиБ№ЪЧЦјКђБфЖЏЃЈШчБљЦкЃЉЕФгАЯьЃЌвђДЫЃЌдкЭЌвЛИіЧјгђЩњЛюЕФЮяжжгаПЩФмЪЧдВњЕФвВгаПЩФмЪЧЧЈвЦЖјРДЕФЁЃЕБШЛЃЌдкЕиЪЗГпЖШЩЯЃЌЙЙдьдЫЖЏЖдЩњЮяЖрбљадЕиРэИёОжЕФгАЯьПЩФмИќЮЊЩюдЖЃЌе§ШчПМПЫЫЙКЭФТЖћЃЈ2007ЃЉжИГіЕФФЧбљЃКЁАЩњЮяЖрбљадЕиРэИёОжЪЧЮвУЧаааЧЩЯНјЛЏКЭАхПщЙЙдьетСНЬЈЮАДѓЛњЦїЯрЛЅзїгУЕФНсЙћЁЁТНПщЦЦЫщКЭдЫЖЏЕМжТгаЙиЕФЩњЮяРрШКБЛдЖИєЃЌЖјХізВАбЦ№дДЪЎЗжВЛЭЌЕФЁЂВЛдјЯыЕНЕФРрШКДјЕНСЫвЛЦ№ЁБЁЃ
ГЄЦквдРДЃЌЮяжжЕФЗжВМИёОжЪЧЩњЮябЇЁЂЩњЬЌбЇЁЂНјЛЏЩњЮябЇЁЂЩњЮяЕиРэбЇЁЂБЃЛЄЩњЮябЇЕШжюЖрЩњУќПЦбЇСьгђЕФПЦбЇМвЫљЙизЂЕФживЊЮЪЬтжЎвЛЁЃЯждкШЫУЧЯраХЃЌЪРНчЩЯЪ§вдАйЭђМЧЕФаЮаЮЩЋЩЋЕФЮяжжЖМЦ№дДздЕЅвЛЕФзцЯШЮяжжЁЃетРяВЛЩюШыЬжТлЮяжжЪЧШчКЮНјЛЏЖјРДЕФЯИНкЃЌЖјжиЕуНщЩмЩњЮяЖрбљадЕФЕиРэИёОжМАЦфЛљБОГЩвђЃЌвдБуДгЧјгђЕиРэГпЖШЩЯРэНтздШЛНчжаЮяжжЕФКъЙлЩњЬЌбЇЩшМЦдРэЃЌЕЋЪЁТдСЫгыжжЯЕЯрЙиЕФЯЕЭГЗжРрбЇЯИНкЁЃ
вЛЁЂЮяжжЁЊЩњУќзюЛљБОЕФЩњДцЕЅдЊ
1.ЮяжжЕФИХФюЁЊвРШЛФбвдЭГвЛЖЈвх
ШЫРрЖдЮяжжЕФШЯЪЖвВОРњСЫЧњелЕФРњГЬЁЃдчдкЖўЧЇЖрФъЧАЃЌЙХЯЃРАЮАДѓЕФембЇМвбЧРяЪПЖрЕТЬсГіЩњЮяПЩбЯИёЧјЗжЮЊВЛЭЌЕФЁАЮяжжЁБЃЌЕЋШЯЮЊЮяжжЪЧгРКуВЛБфЕФЁЃЪЎАЫЪРМЭЃЌШ№ЕфЮАДѓЕФжВЮябЇМвСжФЮЃЈCarolus LinnaeusЃЉЪзДЮвдЩњжГЦїЙйРДНјааЩњЮяЗжРрЃЌЬсГіСЫвЛаЉЮяжжЮЊЭъУРЕФЃЌЖјСэвЛаЉдђЪЧЗТдьЕФЙлЕуЃЌСжФЮЪБДњЕФЮяжжИХФюЕФЬиеїЪЧВЛБфЃЈЁАЩЯЕлЁБДДдьЃЉгыПЭЙлДцдкЃЌ1735ФъСжФЮЗЂБэЁАжВЮяжжжОЃЈSpeciesPlantarumЃЉЁБЃЌЪзДЮЬсГіСЫвдРЖЁЮФРДЮЊЩњЮяУќУћЕФЫЋУћЗЈЃЈЕквЛИіУћзжЪЧЪєЕФУћзжЃЌЕкЖўИіЪЧжжЕФУћзжЃЌЪєУћЮЊУћДЪЃЌжжУћЮЊаЮШнДЪЃЉЃЌДЫжжУќУћЗЈвВвЛжБбггУжСНёЁЃ
ЪЎОХЪРМЭГѕЃЌЗЈЙњжјУћЕФВЉЮябЇМвРТэПЫЃЈJean-Baptiste LamarckЃЉдкЦфжјзїЁАPhilosophie zoologiqueЃЈвыЃКЖЏЮябЇембЇЃЉЁБЃЈdeLamarck 1809ЃЉжаЗёЖЈСЫДДдьТлЃЌШЯЮЊЫцзХЪБМфЕФЭЦвЦЃЌЮяжжНЋж№НЅЗЂЩњБфЛЏЖјаЮГЩаТЮяжжЃЌМДЬсГіСЫВЛЭЌЮяжжжЎМфПЩвдгаСЌајВЛЖЯЕФЯЕеЕФЙлЕуЃЌЫфШЛЫћЮДФме§ШЗЫЕУїЮяжжЕФбнБфЛњжЦЁЃДяЖћЮФЃЈDarwin 1859ЃЉЭЈЙ§БфвьгыздШЛбЁдёЯЕЭГЕиВћЪіСЫЮяжжПЩБфЕФЛњжЦЃЌЕЋЪЧЫћШЯЮЊЮяжжИХФюЪЧШЫЮЊЕФЕЅдЊЃЌвђДЫУЛгаПМТЧЮяжжЕФЖЈвхЁЃДяЖћЮФЕФНјЛЏТлНвЪОСЫЮяжжЦ№дДЕФРњЪЗСЌајадЃЌШДКіЪгСЫЮяжжЕФЯрЖдЮШЖЈЁЃ
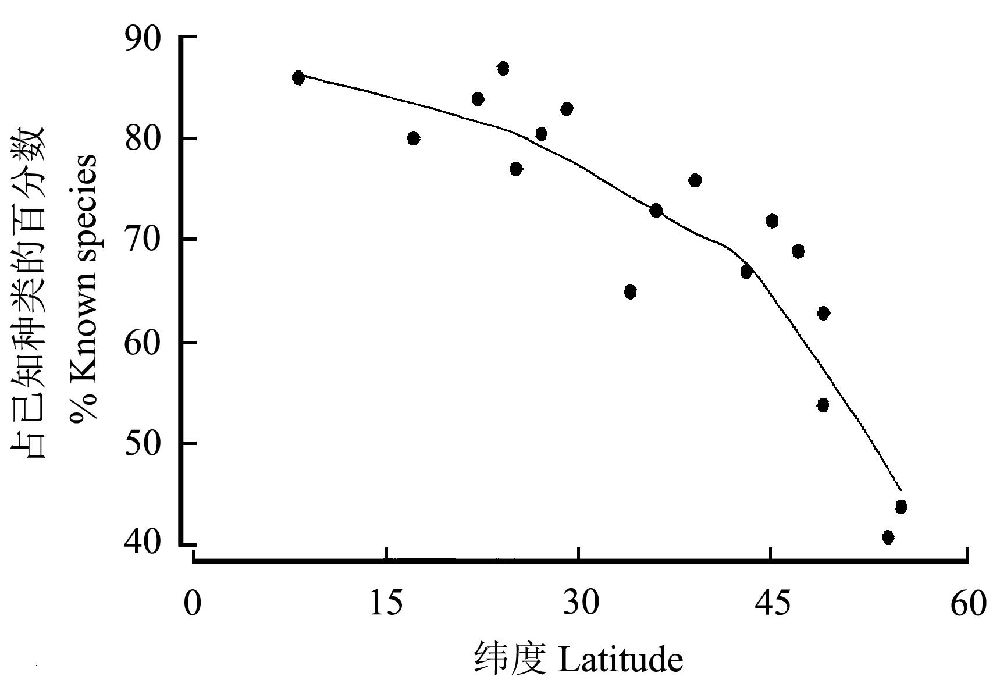
1942ФъЃЌЖїЫЙЬиЁЄТѕЖћЬсГіСЫЩњЮябЇЮяжжИХФюЃКЁАФмЙЛБЫДЫНЛХфЕФздШЛжжШКзщГЩЕФвЛИіРрШКЃЌЦфдкЩњжГЗНУцгыЦфЫћетбљЕФРрШКжЎМфБЫДЫИєРыЁБЃЌетОЭЪЧЫЕЃЌЮяжжЕУвдЧјЗжВЛЪЧДПДтвђЮЊЫќУЧПДЦ№РДВЛвЛбљЃЌЖјЪЧвђЮЊЫќУЧжЎМфДцдкЮоЗЈЯрЛЅНЛХфЕФБкРнЃЌетбљвЛИіЮяжжОЭЪЧвЛИіЗБжГШКЬхЃЌвЛИіЛљвђПтЃЈПЦвђ2009ЃЉЁЃ
ЯжДњЕФЩњЮябЇМвЦеБщШЯЮЊЃЌЮяжжЃЈSpeciesЃЉМШЪЧЩњЮяЗжРрЕФЛљБОЕЅдЊЃЌвВЪЧЩњЮяЩњДцЁЊЗБбмЁЊБфвьЁЊЪЪгІЁЊНјЛЏЕФЛљБОЕЅдЊЁЃЕЋЪЧЃЌгЩгкЩњУќаЮЪНЕФМЋЖЫЖрбљадЃЌМДБуЪЧНёЬьЃЌХаЖЈЮяжжЕФБъзМвВВЛОЁвЛжТЃЌгабЇепЪЙгУаЮЬЌбЇжжЃЌвВгаЪЙгУЩњжГИєРыжжЁЃЖдИпЕШЖЏжВЮяРДЫЕЃЌЩњжГИєРыЪЧХаЖЯЮяжжЕФгааЇБъзМЃЌЕЋЪЧЖдгквЛаЉШБЗІгаадЩњжГЕФЩњЮяЃЈШчЪєгкдКЫЩњЮяЕФЯИОњЕШЃЉЃЌжЛФмвРППаЮЬЌЁЂЩњЛЏМАвХДЋЮяжЪЕШЕФВювьРДХаЖЈЁЃМДБуШчДЫЃЌЮяжжШдШЛЪЧЫљгаЩњУќПЦбЇСьгђзюВЛФмКіЪгЕФИХФюЁЃ
2.ЯжДцЕФЮяжжЁЊвбУшЪі170ЖрЭђжж
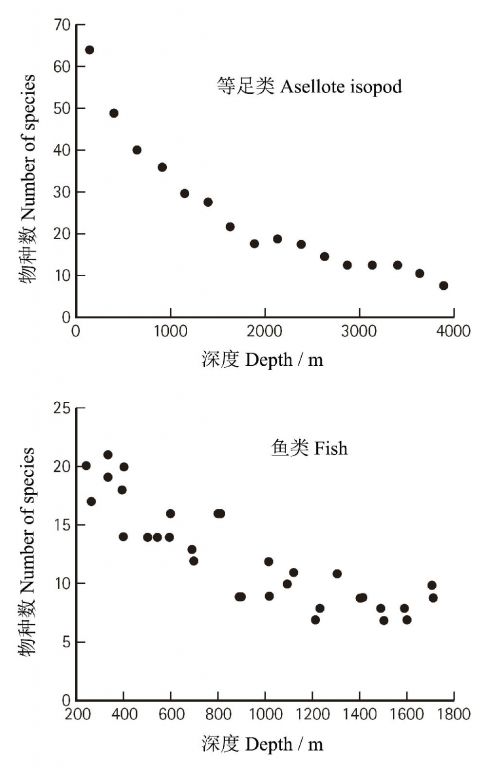
дкНщЩмЮяжжЗжВМЕиРэИёОжжЎЧАЃЌгаБивЊПДПДЕиЧђЩЯЕНЕзгаЖрЩйЮяжжЃПетЮЪЬтПДЫЦМђЕЅЃЌЦфЪЕШДФбвдЛиД№ЁЃОнGroombridge КЭJenkinsЃЈ2002ЃЉЙРМЦЃЌвбУшЪіЕФжжРрдМЮЊ170ЭђжжЃЌЙРМЦЕиЧђЩЯЮяжжЕФжжЪ§ЖрДя1400ЭђжжЃЈБэ1ЃЉЃЁдквбОУшЪіЕФЮяжжжаЃЌгађІЖЏЮяЕФРЅГцКЭЖрзуРрзюЖрЃЈ96.3ЭђжжЃЉЃЌжВЮяДЮжЎЃЈ27ЭђжжЃЉЃЌМЙзЕЖЏЮяВЂВЛЖрЃЈ5.25ЭђжжЃЉЃЌзюЩйЕФРрШКЮЊЙХЩњОњЃЌжЛга175жжЁЃПДРДЃЌзюдЪМЃЈМђЕЅЃЉКЭзюИпЕШЃЈИДдгЃЉЕФЩњЮяРрШКЦфжжРрЪ§ЯрЖдНЯЩйЁЃ
ЫфШЛЯИОњКЭЙХОњзюЮЊЙХРЯЃЌЕЋЦфжжРрЪ§ШДзюЩйЃЌвђЮЊКмФбБЃДцдкЛЏЪЏжаЃЌвВКмФбЯёИпЕШЖЏжВЮяФЧбљЭЈЙ§ЛЏЪЏНјааРњЪЗбнЛЏЕФЗжЮіЁЃгЩгкЯжДцЕФЯИОњПЩеЙЯжОЊШЫЕФБфвьЫйЖШЃЈгыЕижЪФъДњЯрБШЃЉЃЌвђДЫКмФбЯыЯѓЯИОњжжРрЪ§ВЛЖрЪЧгЩгкЦфЗжЛЏЫйЖШТ§ЕФдЕЙЪЁЃФЧУДЃЌвЛжжПЩФмОЭЪЧЯИОњвдИпЫйЕФБфвьЪЪгІЛЗОГЃЌдкМђЕЅЕФЛљвђзщЩЯНјаазХвХДЋНсЙЙЕФПьЫйИќаТЁЃ
Бэ1 ЯждквбжЊЕФЩњЮяжївЊРрШКЕФжжРрЃЌАќРЈЩаЮДБЛЗЂЯжЕФжжРрЕФЙРЫу
ецКЫНч | вбУшЪіЕФжжРрЪ§ | ЙРМЦЕФзмЪ§ | |
ЙХЩњОњ |
| 175 |
|
ЯИОњ |
| 10,000 |
|
ецКЫЩњЮя |
|
|
|
| ЖЏЮяНчAnimalia |
| 1,320,000 |
| МЙзЕЖЏЮяУХЃЈзмМЦЃЉ Craniata (vertebrates), total | 52,500 | 55,000 |
| ВИШщЖЏЮяMammals | 4,600 |
|
| ФёРрBirds | 9,750 |
|
| ХРааЖЏЮяReptiles | 8,002 |
|
| СНЦмЖЏЮяAmphibians | 4,950 |
|
| гуРрFishes | 25,000 |
|
| гаЖѕЖЏЮяЃЈРЅГцКЭЖрзуРрЃЉ Mandibulata (insects and myriapods) | 963,000 | 8,000,000 |
| ђќжЋЖЏЮяЃЈжЉжыЕШЃЉ Chelicerata (arachnids, etc.) | 75,000 | 750,000 |
| ШэЬхЖЏЮяMollusca | 70,000 | 200,000 |
| МзПЧЖЏЮяCrustacea | 40,000 | 150,000 |
| ЯпГцNematoda | 25,000 | 400,000 |
| ецОњFungi | 72,000 | 1,5000,000 |
| жВЮяНчPlantae | 270,000 | 320,000 |
| дЩњЩњЮяProtoctista | 80,000 | 600,000 |
змМЦ |
| 1,750,000 | 14,000,000 |
ЃЈв§здGroombridge and Jenkins 2002ЃЉ
3.ЮяжжЕФЗжЛЏЁЊЪмжЦгкЩњДцЬѕМўЕФЫцЛњбнЛЏ
ЮяжжЪЧдѕбљаЮГЩЕФЃПДяЖћЮФШЯЮЊЫљгаЮяжжЖМЪЧбнЛЏЕФВњЮяЃЌЖјБфвьгыздШЛбЁдёЪЧетвЛЙ§ГЬЕФжївЊЭЦЖЏСІЁЃаТЮяжжЕФаЮГЩЙ§ГЬвВГЦЮЊЮяжжЗжЛЏЃЌжївЊАќРЈвьгђЗжЛЏЃЈallopatric speciationЃЉКЭЭЌгђЗжЛЏЃЈsympatric speciationЃЉЃЌЖјБпдЕЗжЛЏЃЈperipatricЃЉКЭСйгђЗжЛЏЃЈparapatric speciationЃЉЫЦКѕЮЊЙ§ЖЩРраЭЁЃ
вьгђЗжЛЏжИЭЌвЛЮяжжгЩгкЕиРэИєРыЃЌЗжБ№бнЛЏЮЊВЛЭЌЮяжжЕФЙ§ГЬЁЃетОЭЪЧЖїЫЙЬиЁЄТѕЖћЕФЙлЕуЁЃжжШКИєРыПЩЭЈЙ§ЕижЪЛюЖЏЛђЦјКђБфЛЏЃЈШчДѓТНЦЏвЦЁЂЩНТіЛђЯПЙШЕФаЮГЩЁЂБљДЈРЉеЙЛђЮЎЫѕЁЂТНЧХЕФаЮГЩЛђЯћЪЇЁЂЩГФЎЕФРЉДѓЛђЫѕаЁЕШЃЉРДЪЕЯжЃЌвВПЩЭЈЙ§жжШКЕФРЉЩЂРДЪЕЯжЃЌЦЉШчЖЏЮяЕФЧЈсуЛђвтЭтЕФЧЈвЦЖМПЩФмЗжИюжжШКЃЌШчЙћСНИіжжШКжЎМфЭЃжЙСЫЛљвђНЛСїЃЌОЭПЩФмЕМжТаТЮяжжЕФаЮГЩЁЃЕБШЛЃЌИєРывЛАуЪЧЕиРэЩЯЕФИєРыЃЌЕЋВЛвЛЖЈЖМашвЊЕиРэБкРнЁЃЦфЪЕЃЌДяЖћЮФдкЦфЁАЮяжжЦ№дДЁБвЛЪщжаОЭвбОзЂвтЕНЕиРэвђЫидкЮяжжЗжЛЏжаЕФживЊзїгУЁЃвьгђЗжЛЏЕФР§згЪ§ВЛЪЄЪ§ЃЌЦЉШчЃЌДѓамУЈЪЧЪєгкЪГШтФПЕФвЛжжамЃЌИУФПжаДѓЖрЪ§ГЃМћЕФамЖМЪЧдгЪГадЖЏЮяЃЌЕЋамУЈМИКѕжЛГджёзгЃЌЫќЩњЛюдкжаЙњЮїВПИпКЃАЮЩНЧјЕФУЏУмжёСжжаЃЌФЧРяУЛгаВЖЪГепЕФЧжШХЃЌЫќУЧУПЬьЛЈ10-12аЁЪБНРЪГжёзгЃЈЙХЖћЕТ2008ЃЉЁЃ
ЭЌгђЗжЛЏжИЭЌвЛЮяжждкЯрЭЌЕФЛЗОГЃЌгЩгкааЮЊЯАадЕФИФБфЛђЛљвђЭЛБфЕШдвђЖјбнЛЏГЩВЛЭЌЮяжжЕФЙ§ГЬЁЃетЦфЪЕЪЧДяЖћЮФЙлЕуЕФЯжДњУшЪіЃЌДяЖћЮФШЯЮЊЃЌжЛвЊгаЩњДцОКељЃЌдкаэЖрЕиЧјЖМПЩФмВњЩњаТЮяжжЁЃдкЮЛгкЖЋЗЧДѓСбЙШЕФТэРЮЌКўЃЈLake MalawiЃЉжаЃЌРігуЃЈcichlidfishesЃЉЕФЮяжжЗжЛЏОЭЪЧЭЌгђЗжЛЏЕФКмКУР§жЄЃЌдкЙ§ШЅЕФ70ЭђФъЦкМфЃЌДгЕЅвЛЕФРігузцЯШЗжЛЏГіГЌЙ§400ИіЮяжжЃЈDanley & Kocher 2001ЃЉЁЃ
ЮЊЪВУДвЊВЛЖЯаЮГЩаТЮяжжЃПДяЖћЮФШЯЮЊЮяжжЪЧЮЊСЫЬюВЙФГжжЩњЬЌЮЛЕФПеШБВХГіЯждкДѓздШЛжаЃЌЖјгабЇепГжЭъШЋВЛЭЌЕФПДЗЈЃЌШчПЦвђЃЈ2009ЃЉШЯЮЊЃЌВЛЪЧвђЮЊздШЛНчашвЊВЛЭЌЕФЮяжжЃЌЮвУЧВХгЕгаСЫВЛЭЌЕФЮяжжЃЌЮяжжжЛЪЧбнЛЏЕФвтЭтЃЌЁАВЛЭЌРрШКЁБЖдЩњЮяЖрбљадЖјбдКмживЊЃЌЕЋЫќУЧВЂВЛЪЧвђЮЊдіМгСЫЩњЮяЖрбљадВХБЛбнЛЏГіРДЃЌвВВЛЪЧвђЮЊЬсЙЉСЫЦНКтЕФЩњЬЌЯЕЭГВХБЛбнЛЏГіРДЃЌЫќУЧНіНіЪЧдкПеМфЩЯБЫДЫИєРыЕФжжШКЯђзХВЛЭЌЗНЯђбнЛЏЫљВњЩњЕФЛљвђБкРндьГЩЕФВЛПЩБмУтЕФБиШЛНсЙћЖјвбЁЃЕЋПЦвђЕФетжжПДЗЈЯдШЛЮоЗЈНтЪЭЯёдкТэРЮЌКўжаРігуЕФПьЫйЗжЛЏЃЈЭЌгђЗжЛЏЃЉЯжЯѓЁЃ
вРЮвПДРДЃЌВЛБиНЋетСНжжЙлФюЖдСЂЦ№РДЁЃЮяжжЕУвдЗжЛЏЃЌДгЩњЬЌбЇНЧЖШРДПДБиаыгаПеШБЕФЩњЬЌЮЛПЩЬюЃЌЕЋЪЧЃЌЮяжжЮЊКЮдквЛаЉЕиЗНЗжЛЏПьЖјдкСэвЛаЉЕиЗНЗжЛЏТ§ЃЌВЂВЛЭъШЋЪЧвЛжжЫцЛњЕФНсЙћЃЌвВЮоЗЈЭъШЋгУИєРыГЬЖШРДНтЪЭЃЌЯрЗДПЩФмдкКмДѓГЬЖШЩЯШЁОігкЮяжжЕФЩњДцЬѕМўЃЈАќКЌЩњЬЌЮЛЃЉЁЃетОЭЪЧБОеТвЊЬНЬжЕФКЫаФЮЪЬтЁЃ
ЖўЁЂЮяжжЖрбљадЕиРэИёОжЁЊТНЩњжВЮя
1.ЮГЖШаЇгІЁЊЮГЖШдНЕЭЃЌЮяжждНЗсИЛ
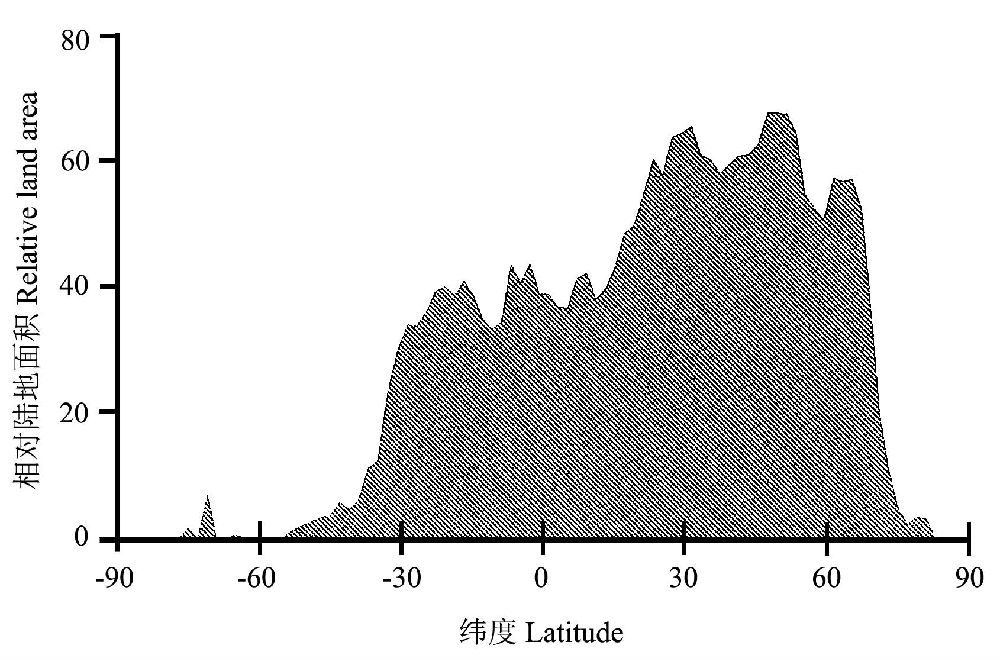
дчдкЪЎОХЪРМЭГѕЃЌХЗжоЕФВЉЮябЇМвЃЈHumboldt and Bonpland 1807ЃЉОЭЙизЂЕНЮяжжЖрбљадбиЮГЖШЬнЖШЕФБфЛЏЧїЪЦЁЃНќДњЕФбаОПБэУїЃЌЪїжжЖрбљадбиЮГЖШЬнЖШЕФИёОжЪЎЗжУїЯдЃЌЭЈГЃЪЧДгИпЮГЖШЯђГрЕРГЪжИЪ§діГЄЃЌЖјЧвдкББАыЧђЯђИпЮГЖШЕиЧјЕФЗжВМЗЖЮЇИќЙувЛаЉЃЈЭМ1ЁЂЭМ2ЃЉЁЃетжжЧПСвЕФЮГЖШаЇгІОйР§ЫЕУїЃЌШчЭЌбљУцЛ§ЕФвЛЦЌЪїСжЃЌдкЖэТоЫЙЖЋФЯВПЃЈ45oNЃЉжЛга40жжЪїЃЌЖјдкТэРДЮїбЧАыЕКЃЈ1oNЃЉШДга1400жжЪїЃЌНЋЫљгажВЮяКЯВЂдквЛЦ№вВгаетжжЧїЪЦЃЈAdams 1988ЃЌ2009ЃЉЁЃвЛАуШЯЮЊЃЌЮГЖШЬнЖШЩЯЕФЮТЖШВювьгІИУЪЧгАЯьЪїжжЖрбљадЕиРэИёОжЕФжївЊвђЫиЁЃ
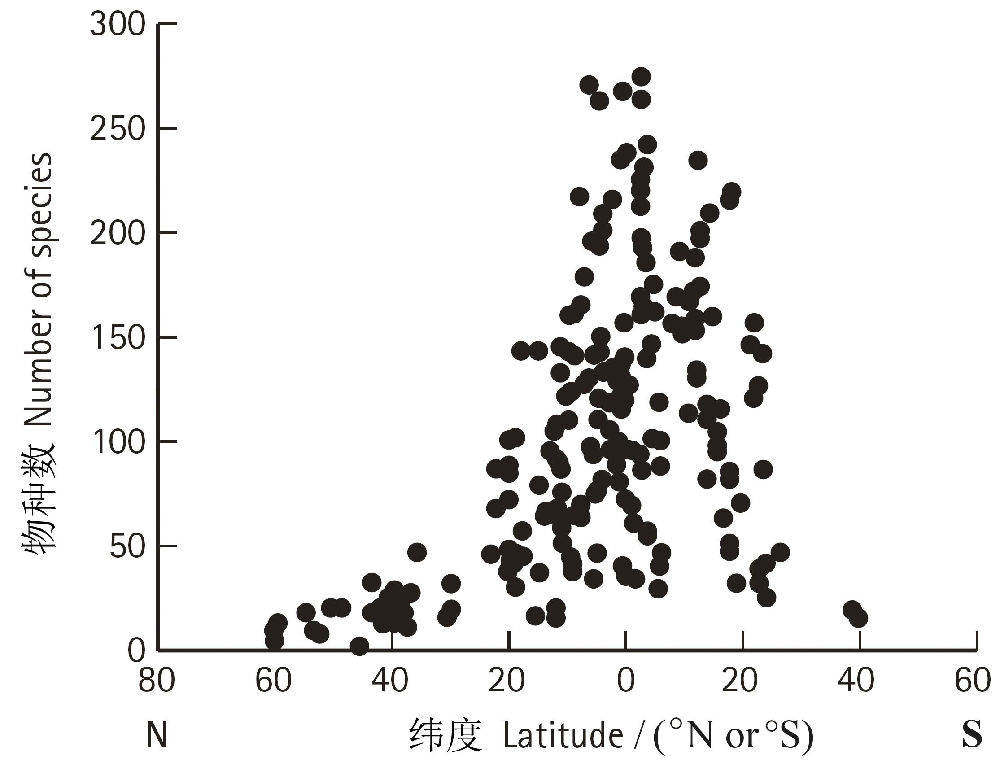
ЭМ1 ШЋЧђЪїжжЖрбљадЃЈбљЗНДѓаЁЁЊ0.1 haЃЉЕФЮГЖШЬнЖШЃЈЗТEnquist& Niklas 2001ЃЉ

ЭМ2 ЖЋбЧЕФвАЩњЪїжжЃЈв§здAdams 2009ЃЉ
ЕЋЪЧЃЌдквЛаЉЧщПіЯТЃЌЮяжжЖрбљадбиОЖШЃЈЖЋЁЊЮїЃЉЗНЯђвВГЪЯжУїЯдЕФЬнЖШЃЌетдкЕЭЮГЖШЕФШШДјЁЂбЧШШДјЕиЧјгШЦфУїЯдЃЌМДЫфШЛЭЌбљЪЧЮТХЏЕФЦјКђЃЌЕЋЪЧгЩгкНЕгъСПЕФОоДѓБфЛЏЃЌЪЊШѓЖШБфЛЏКмДѓЁЃгЩгкЫЎЗжвВЪЧгАЯьжВЮяЩњГЄЕФживЊвђЫиЃЌЯрЫЦЮТЖШЬѕМўЯТЕФЫЎЗжЬнЖШвВОЭЕМжТСЫжВБЛИёОжвђЖјЮяжжЖрбљадИёОжЕФЯджјБфЛЏЃЈЭМ3ЃЉЁЃЦЉШчЃЌДгУРЙњЖЋВПЪЊШѓЕФЦјКђЁњжаВПИЩдяЕФДѓЦНдЧјгђЁњФЯВПЩГФЎЃЌЪїжжЕФЖрбљадУїЯдЯТНЕЃЈCurrie & Paquin 1987, Adams 2009ЃЉ

ЭМ3 ББУРЕФЪїЮяжжЗсЖШЭМЃЈв§здAdams 2009ЃЉ
2.ЮТЖШаЇгІЁЊЮТЖШдНИпЃЌЮяжждНЗсИЛ
ЮТЖШЭљЭљЪЧжВЮяЗжВМЕФЙиМќвЊЫиЃЌвђДЫЃЌвВгАЯьЕНжВЮяЮяжжЖрбљадЕФЕиРэИёОжЁЃЭМ4ЪЧЯдЛЈжВЮяПЦЕФЪїФОгызюЕЭЮТЖШжЎМфЕФЙиЯЕЃЌГЪЯжГіКмКУЕФжБЯпе§ЯрЙиЙиЯЕЃЌМДзюЕЭЮТЖШдНЕЭЃЌПЦЕФЪ§ФПдНЩйЃЌЖјзюЕЭЮТЖШдНИпЃЌПЦЕФЪїФОдНЖрЃЌЫфШЛФЯББАыЧђжЎМфДцдквЛЖЈЕФВювьЃЈПЩФмгыДѓТНИєРыЕФРњЪЗгаЙиЃЉЁЃ
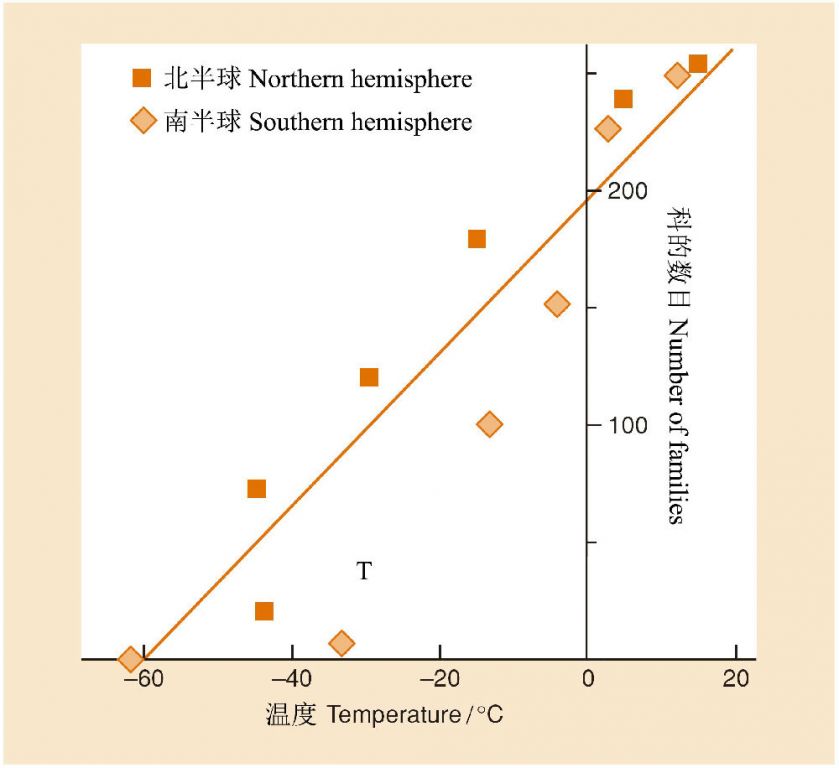
ЭМ4 ФЯББАыЧђЯдЛЈжВЮяПЦЕФЪ§ФПгызюЕЭЮТЖШжЎМфЕФЙиЯЕЃЈBegon et al. 2006в§зд Woodward 1987ЃЉ
3.КЃАЮаЇгІЁЊдкИпКЃАЮЕиЧјЃЌЮяжжЪ§МБОчЯТНЕ
Grytnes & VetaasЃЈ2002ЃЉНЋФсВДЖћЯВТэРбХЩНТі6000mЕФКЃАЮИпЖШвд100-mМфИєЗжЮЊ60ИіДјЃЌБШНЯСЫЯдЛЈжВЮяЕФжжРрЪ§ЫцКЃАЮИпЖШЕФБфЖЏЁЃдкФсВДЖћФЯВПИпдвЛжБЕНКЃАЮ1000mзѓгвЮЊШШДјЦјКђЃЌдк1000-2000mжЎМфЮЊбЧШШДјЛђХЏЮТДјЦјКђЃЌ2000-3000mжЎМфЮЊКЎЮТДјЦјКђЃЌ4000-4500mЮЊбЧИпЩНДјЕФЩЯЯоЃЌвВЪЧЪїФОЯпЃЛдкИпЩНДјЃЌЗжВМгавЛаЉЙрФОЃЌЕЋВнЕиеМгХЪЦЃЌЭљИќИпЕФЕиЗНЃЌжВБЛИќМгВЛСЌајЃЈDobremez 1976ЃЌGrytnes& Vetaas 2002ЃЉЁЃдкКЃАЮИпЖШ1500mвдФкЃЌЮяжжЪ§ЫцКЃАЮИпЖШЕФдіМгЖјПьЫйдіМгЃЌдк1500-2500mжЎМфЃЌжжРрЪ§КмЩйБфЛЏЃЌГЌЙ§етвЛИпЖШЃЌЮяжжЪ§ЮШВНЯТНЕЃЈЭМ5ЃЉЁЃжЕЕУзЂвтЕФЪЧЃЌдк0-100mжЎМфЃЌНіга62ИіЮяжжБЛМЧТМЃЌетПЩФмЪЧгЩгкдкФсВДЖћетаЉЕЭЮГЖШЧјгђЕФУцЛ§КмаЁЕФдЕЙЪЁЃ

ЭМ5 ФсЦШЖћЯВТэРбХЩНТіЯдЛЈжВЮяЕФжжРрЪ§ЫцКЃАЮИпЖШЕФБфЖЏЃЈЗТGrytnes& Vetaas 2002ЃЉ
4.НЕгъСПаЇгІЁЊНЕгъСПдНДѓЃЌЮяжждНЗсИЛ
OЁЏBrienЃЈ1993ЃЉбаОПСЫЗЧжоФЯВПФОБОжВЮяЃЈN=1372жжЃЉЖрбљадЕФЗжВМИёОжЃЌЮяжжЖрбљадГЪЯжГіДгЮїЯђЖЋж№НЅдіМгЕФЧїЪЦЃЌдкИЩКЕАыИЩКЕЕиЧјзюЕЭЃЌдкЪЊШѓЕиЧјзюИпЃЌгыНЕгъСПГЪЯжГіУїЯдЕФе§ЯрЙиЙиЯЕЃЈЭМ6ЃЉЁЃ

ЭМ6 ЗЧжоФЯВПФОБОжВЮяЖрбљадгыНЕгъСПжЎМфЕФЙиЯЕЃЌЭјИёУцЛ§ЮЊ20,000 km2ЃЈЗТOЁЏBrien 1993ЃЉ
Currie & PaquinЃЈ1987ЃЉбаОПСЫББУРЪїжжЕФЗжВМЃЌНЋУРЙњКЭМгФУДѓЧјЗжЮЊ336ИібљПщЃЌЙВЭГМЦСЫ620ИіЭСОгЪїжжЃЈИпгк3mЕФжВЮяЃЉЁЃЭЈЙ§НЋЪїжжРрЪ§гыЛЗОГвђзгНјааЯрЙиЗжЮіЗЂЯжЃЌдкИїжжЛЗОГвђзгжаЃЌгыЭСШРЫЎЗжеєЗЂеєЬкЫ№ЪЇзмСПЃЈevapotranspirationЃЌМДДгЭСШРеєЗЂЛђДгжВЮяеєЬкЕФзмЫЎЗжСПЃЉЕФЯрЙиадзюКУЃЈЭМ7ЃЉЁЃЫфШЛЫћУЧгУТпМЫЙЕйЧњЯпНќЫЦЃЌЕЋЪЕМЪвВШБЗІЧїЯђБЅКЭЕФНЅНќЯпЧјгђЃЌЖдЭМ7ЦфЪЕвВПЩвдгУжБЯпУшЪіЃЌЩѕжСвВФмгУжИЪ§КЏЪ§РДУшЪіЁЃ
ашвЊжИГіЕФЪЧЃЌВЛФбЗЂЯжЃЌдкАыЩГФЎКЭЩГФЎЕиЧјЃЌеєЗЂСПКмИпЃЌЕЋгЩгкНЕгъСПЩйЃЌЦфЪЕжВЮяЬиБ№ЪЧЪїЪЧКмФбЩњГЄЕФЃЌЪ§жжЖрбљадВЛПЩФмИпЃЌвђДЫЃЌдкЬНЬжгыЪїжжЖрбљадЪБЃЌПЩФмгУЪЊШѓЖШИќМгКЯРэЁЃ

ЭМ7 НЋЪїжжЖрбљадзїЮЊЭСШРЫЎЗжеєЗЂеєЬкЫ№ЪЇФъзмСПЕФКЏЪ§ЃЈn=366ЃЉЃЌЪЕЯпЮЊФтКЯЕФТпМЫЙЕйФЃаЭЃЈв§здCurrie & Paquin 1987ЃЉ
5.ЩњВњСІаЇгІЁЊЩњВњСІдНИпЃЌЮяжжЪ§дНЖр
дкЮТДјЩСжЧјЃЌЪїжжЖрбљадЫцОЛГѕМЖЩњВњСІЃЈжївЊвРРЕгкЮТЖШЃЉЕФдіМгЖјдіМгЃЈЭМ8ЃЉЃЌЕЋЪЧвВДцдквЛЖЈЕФЧјгђВювьЃКгыББУРЯрБШЃЌбЧжоЕиЧјНЯЮТХЏЕФЦјКђЬѕМўЯТЕФЮяжжИёЭтЕиЗсИЛЃЌЦЉШчЃЌЭЌбљЕФЮТДјЕиЧјЃЌзюЮТХЏГБЪЊЕФФЯХЗЃЈШчАЭЖћИЩЃЉКЭББУРЃЈШчЧЧжЮбЧФЯВПЁЂМгРћИЃРћбЧФЯВПЁЂИЅТоРяДяББВПЃЉгыбЧжоЭЌбљЕФЮЛжУЃЈШчШеБОФЯВПКЭЬЈЭхЃЉЯрБШЃЌЪїЕФжжРрвЊЩйЕУЖрЃЌетПЩФмгыбЧжоЕФЪїжждкБљЦкИќКУЕиДцЛюЯТРДСЫгавЛЖЈЙиЯЕЃЈAdams 2009ЃЉЁЃ

ЭМ8 ГѕМЖЩњВњСІКЭЪїЮяжжЖрбљадЕФЙиЯЕЁЃЁјЃКЪ§ОнРДздЖЋбЧЃЌЁѕУРЙњЖЋББВПЃЈв§здAdam 2009ЃЉ
Ш§ЁЂЮяжжЖрбљадЕиРэИёОжЁЊТНЩњЖЏЮя
1.ЮГЖШаЇгІЁЊДгЕЭЮГЕНИпЮГЃЌЮяжжЪ§ж№НЅНЕЕЭ
ЃЈ1ЃЉКуЮТЖЏЮяЁЊФёРрКЭВИШщЖЏЮя
ЭЈЙ§ЖдУРжоББВПКЭжаВПЕиЧјЕФЗБжГФёРржжЪ§ЕФбаОПЗЂЯжЃЌФёРржжЪ§ДгИпЮГЖШЯђЕЭЮГЖШЗНЯђГЪЯжУїЯдЕФжИЪ§діГЄЙиЯЕЃЈЭМ9ЃЉЃЌМДдкИпЮГЖШЕиЧјжЛга20-30жжФёЃЌЖјдкГрЕРИННќЃЌФёРржжЪ§ГЌЙ§1000жжЃЁ
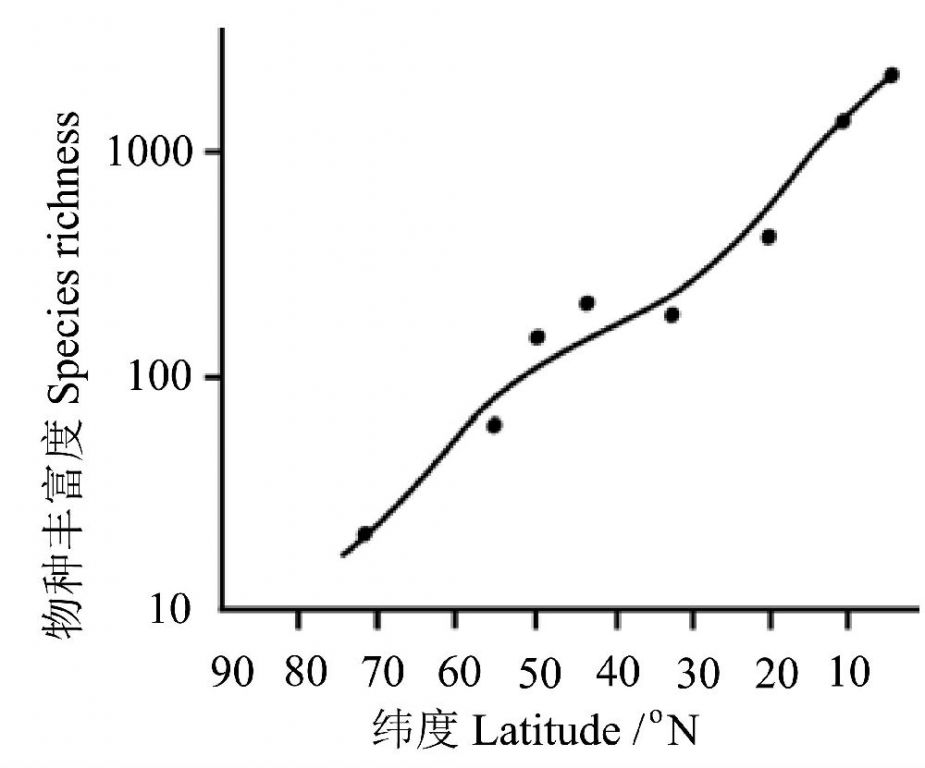
ЭМ9 УРжоББВПКЭжаВПЕиЧјЕФЗБжГФёРрЮяжжЗсИЛЖШППНќГрЕРзюИпЃЈв§здHuston 1993ЃЉ
ЭЈЙ§ЖдПчдНаТДѓТНЕФФёРрЖрбљадбиЮГЖШЬнЖШЕФЗжВМИёОжЕФЗжЮіЗЂЯжЃЌдкФЯББАыЧђОљЪЧЫцЮГЖШЕФдіМгФёРрЮяжжЪ§ЯТНЕЃЌЧвдкЭЌбљЕФЮГЖШЃЌФЯАыЧђЕФФёРржжРрЪ§ТдИпгкББАыЧђЃЈЭМ10ЃЉЁЃ

ЭМ10 ПчдНаТДѓТНЕФФёРрЖрбљадЕФЮГЖШЬнЖШЃЌЭјИёДѓаЁдМЮЊ611,000 km2ЃЈЗТGaston & Blackburn 2000ЃЉ
аТДѓТНЕФВИШщЖЏЮявВГЪЯжГігыФёРрРрЫЦЕФЮГЖШЬнЖШЗжВМЃЌМДВИШщЖЏЮяЮяжжЖрбљадЫцЮГЖШЕФЩЯЩ§ЖјЯТНЕЃЌФЯББАыЧђвВгавЛЖЈЕФВювьЃЌзюИпЕФЮяжжЖрбљадГіЯждкФЯАыЧђ0-15ЁужЎМфЃЈЭМ11ЃЉЁЃ

ЭМ11 ПчдНаТДѓТНЕФВИШщРрЖрбљадЕФЮГЖШЬнЖШЃЌЮГЖШМфИєЮЊ2.5ЁуЃЈЗТKaufman& Willig 1998ЃЌGaston & Spice 2004ЃЉ
ЃЈ2ЃЉБфЮТЖЏЮя
ЮоТлдкФЯАыЧђЛЙЪЧББАыЧђЃЌДгИпЮГЖШЭљЕЭЮГЖШЗНЯђЃЌШ§РрБфЮТМЙзЕЖЏЮяЁЊСНЦмРрЁЂђсђцЃЈЪєХРааЖЏЮяЃЉКЭЩпЃЈЪєХРааЖЏЮяЃЉЕФЪєЖрбљадОљГЪРрЫЦгкжИЪ§ЕФдіГЄФЃЪНЃЈЭМ12ЃЉЃЌЫфШЛЫќУЧЕФОјЖдЪєЪ§ДцдкУїЯдЕФВювьЃЌМДЩп>ђсђц>СНЦмРрЁЃ
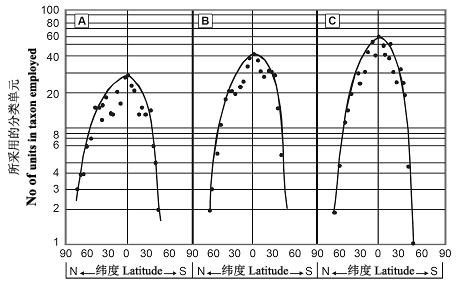
ЭМ12 Ш§ИіБфЮТЖЏЮяРрШКЖрбљадгыЮГЖШжЎМфЕФЙиЯЕЭМЁЃЃЈAЃЉСНЦмРрЕФЪєЪ§ЃЌЃЈBЃЉђсђцЕФЪєЪ§ЃЌЃЈCЃЉЩпЕФЪєЪ§ЃЈв§здMEERЃЌhttp://www.meer.org/ebook/chap3.htmЃЉ
2.ЮТЖШаЇгІЁЊЮТЖШдНЕЭЃЌЮяжждНЦЖёЄ
ЃЈ1ЃЉФёРр
LennonЕШЃЈ2000ЃЉбаОПСЫгЂЙњФёРргыЯФМОдТЦНОљЮТЖШжЎМфЕФЙиЯЕЃЌРћгУСЫгЩгЂЙњФёРрбЇЛљН№зщжЏЕФ2ДЮЖРСЂЕФЕїВщЪ§ОнЃЌвЛДЮЪЧдк1981-83ФъЦкМфНјааЕФЖЌМОЕїВщЃЌСэвЛДЮЪЧ1988-91ФъЦкМфзщжЏЕФЯФМОЕїВщЃЌзмЙВга2362ИібљЗНЃЈУПИі10 x10 kmЃЉЃЌКИЧСЫгЂЙњДѓВПЗжЧјгђвдМАЙуЗКЕФЩњОГЁЂЕиУВКЭЦјКђРраЭЁЃЫћУЧИљОнФёРрЕФЧЈсузДЬЌЃЈСєФёЛђКђФёЃЉКЭМОНкЃЈЯФМОЛђЖЌМОЗжВМЃЉЖЈвхСЫ4жжРраЭЃЌЯФМОЕФСєФёЃЈSRЃЉЁЂЯФМОЕФКђФёЃЈSVЃЉЁЂЖЌМОЕФСєФёЃЈWRЃЉКЭЖЌМОЕФКђФёЃЈWVЃЉЁЃГ§СЫЯФМОЕФКђФёЭтЃЌФёРрЮяжжЖрбљадгыЯФМОЮТЖШжЎМфГЪЯжГіЧПСвЕФе§ЯрЙиЙиЯЕЃЌМДСєФёгыЖЌМОКђФёгыЯФМОЮТЖШжЎМфДцдкУїЯдЕФЯрЙиЙиЯЕЃЈЭМ13ЃЉЁЃвВОЭЪЧЫЕЯФМОЮТЖШзмЬхЩЯРДПДУїЯдгАЯьЕНФёРрЕФЮяжжЖрбљадЁЃ
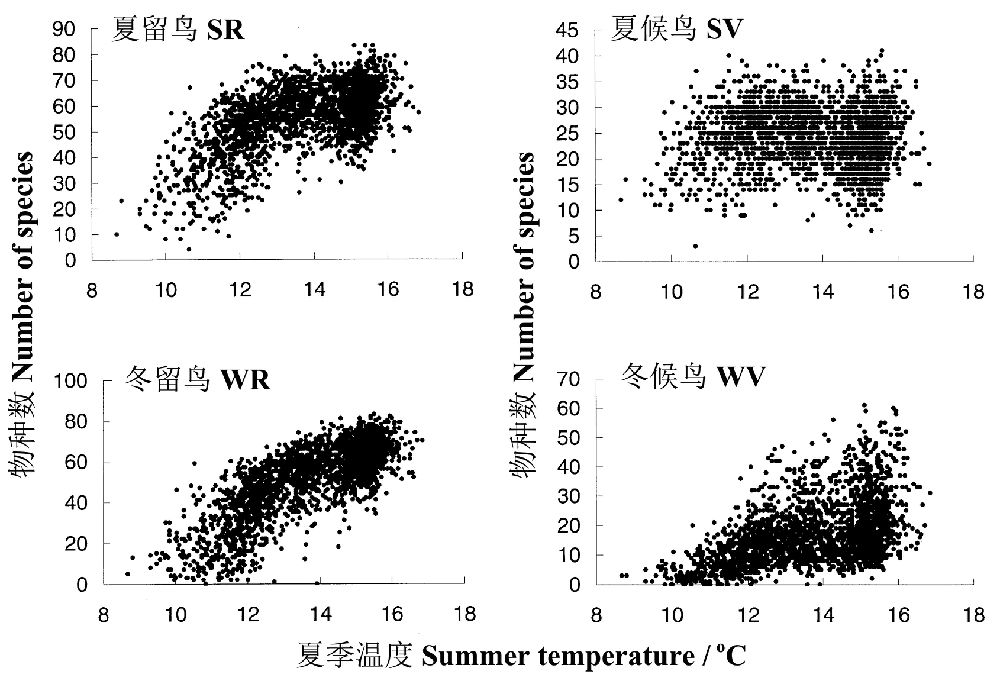
ЭМ13 гЂЙњФёРржжРрЖрбљадЃЈбљЗНЭјИёДѓаЁ10 km x 10 kmЃЉгыдТЦНОљЯФМОЮТЖШжЎМфЕФЙиЯЕЃЈв§здLennon et al. 2000ЃЉ
ЃЈ2ЃЉВИШщЖЏЮя
ЭЈЙ§ЖдПчдНФЯББУРжоЕФ1255ИіЭЌбљУцЛ§ЕФбљЗНжаВИШщЖЏЮяЮяжжЪ§ЃЈЙВАќРЈ1755ИіЮяжжЃЉгыЛЗОГвђзгжЎМфЙиЯЕЕФбаОПЗЂЯжЃЌЕЅЮЛбљЗНЕФЮяжжЪ§гыФъзюЕЭЮТЖШжЎМфДцдкНЯКУЕФе§ЯрЙиЙиЯЕЃЈНќЫЦжИЪ§діГЄЃЉЃЈЭМ14ЃЉЃЌФъзюЕЭЮТЖШЪЧВИШщЖЏЮяЕЅЮЛбљЗНЮяжжЪ§ЕФзюКУЕФдЄВтБфСПЁЃ

ЭМ14 ББУРжоКЭФЯУРжоЕЅЮЛбљЗНДѓТНВИШщЖЏЮяЖрбљадгыФъзюЕЭЮТЖШжЎМфЕФЙиЯЕЃЌЪ§ОнРДзд1755жжВИШщЖЏЮяЃЈв§здMayand McLean 2007ЃЉ
3.КЃАЮаЇгІЁЊдкИпКЃАЮЕиЧјЃЌЮяжжЪ§МБОчЯТНЕ
PattersonЕШЃЈ1998ЃЉбаОПСЫУиТГФЯВПЕФАВЕиЫЙЩНФёРрКЭђљђ№ЃЈђљђ№ЪЧЮЈвЛФмЗЩЯшЕФВИШщЖЏЮяЃЉЮяжжбиКЃАЮЬнЖШЕФЗжВМИёОжЃЌбљЦЗШЁздТэХЌЙњМвЙЋдАКЭЩњЮяШІБЃЛЄЧјЃЌАќРЈСЫ901жжФёКЭ129жжђљђ№ЃЌКмУїЯдЃЌФёРрКЭђљђ№ЕФЮяжжЖрбљадЫцКЃАЮИпЖШЕФЩЯЩ§ЖјЮШВНЯТНЕЃЌЧвЖўРрЩњЮяЕФЮяжжЪ§ЫцКЃАЮЩ§ИпЖјЯТНЕЕФЧїЪЦЪЎЗжРрЫЦЃЈЭМ15ЃЉЁЃЫфШЛЫћУЧвВЕїВщСЫаЁЪѓЃЌЕЋжЛга28жжЃЌЦфЮяжжЪ§гыКЃАЮИпЖШжЎМфЙиЯЕЕФНсТлгІИУЪЎЗжЩїжиЁЃ

ЭМ15 УиТГТэХЌЙњМвЙЋдАКЭЩњЮяШІБЃЛЄЧјжаЕФФёРрКЭђљђ№ЕФЮяжжЪ§КЭКЃАЮИпЖШжЎМфЕФЙиЯЕЃЈв§здPatterson et al. 1998ЃЉ
змЬхЩЯРДЫЕЃЌТНЩњЖЏЮяЕФЮяжжЪ§ЫцКЃАЮИпЖШЕФЩ§ИпЖјНЕЕЭЃЌЕЋЪЧвВгавЛаЉР§ЭтЃЌЦЉШчЃЌПЦТоРЖржнЕФТьвЯжжРрЪ§ЫцКЃАЮИпЖШГЪЯжЭеЗхаЭЕФБфЛЏЃЌМДзюГѕЮяжжЪ§ЫцКЃАЮИпЖШЕФдіМгЖјЩЯЩ§ЃЌдкНгНќ2000mДІДяЕНзюИпЃЌжЎКѓгжж№НЅЯТНЕЃЌЕБШЛЃЌдкИпКЃАЮЕиЧјвВБШЕЭКЃАЮЕиЧјЕФЮяжжЖрбљадЕЭЕУЖрЃЈЭМ16ЃЉЁЃ

ЭМ16 ПЦТоРЖржнЕФТьвЯжжРрЪ§ЫцКЃАЮИпЖШЕФБфЛЏЃЈЗТSanders 2002ЃЉ
4.ЪЊЖШаЇгІЁЊжаЕШЫЎЗжеєЗЂеєЬкСПЕФЕиЧјЃЌЮяжжзюЮЊЗсИЛ
HawkinsЕШЃЈ2003ЃЉЗжЮіСЫЧБдкЭСШРЫЎЗжеєЗЂеєЬкСПЃЈPETЃЉгыЙХББЧјКЭаТББЧјТНЩњФёРрКЭКћЕћЕФЮяжжЖрбљаджЎМфЕФЙиЯЕЃЈЭМ17ЃЉЃЌЗЂЯждкЕЭPETЧјгђЃЌФёРрКЭКћЕћЕФЮяжжЖрбљадОљЫцPETЕФдіМгЖјдіМгЃЌЖјЕБPETГЌЙ§вЛЖЈЕФуажЕЃЌЮяжжЖрбљадПЊЪМЯТНЕЃЈЙХББЧјЃЉЛђВЛдйдіГЄЃЈаТББЧјЃЉЁЃ
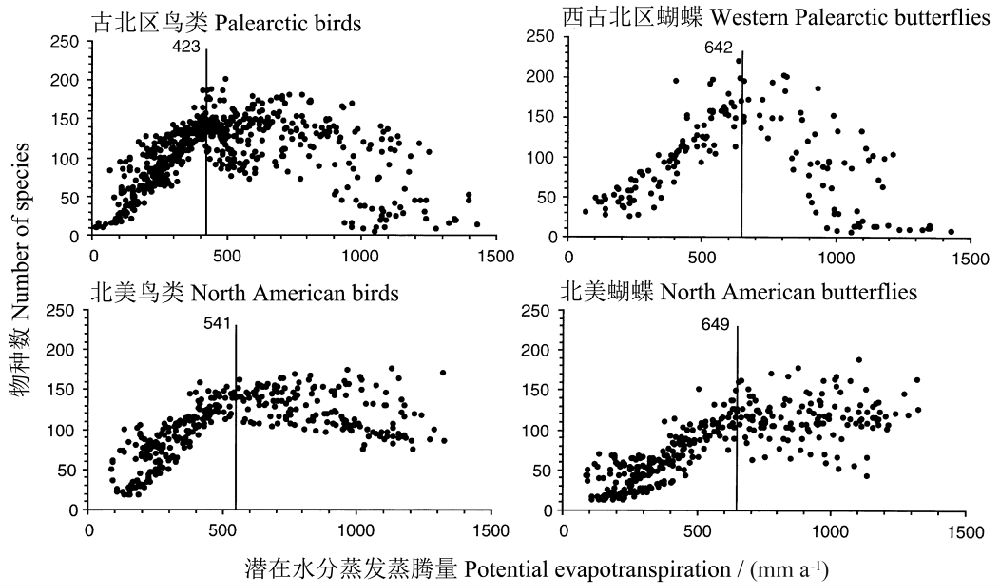
ЭМ17 ЧБдкЭСШРЫЎЗжеєЗЂеєЬкСПЃЈPETЃЉгыЙХББЧјКЭаТББЧјТНЩњФёРрКЭКћЕћЕФЮяжжЖрбљаджЎМфЕФЙиЯЕЁЃДЙЯпБэЪОДге§ЕНИКЃЈЙХББЧјЃЉЛђЕНСуЃЈаТББЧјЃЉЕФзЊелЕуЃЈв§здHawkins et al. 2003ЃЉ
ЫФЁЂЮяжжЖрбљадЕиРэИёОжЁЊЫЎЩњЖЏЮя
1.ЮГЖШаЇгІЁЊдкИпЮГЖШЕиЧјЃЌЮяжжЪ§МБОчЯТНЕ
RoyЕШЃЈ1998ЃЉЪеМЏСЫдкЖЋЬЋЦНбѓКЭЮїДѓЮїбѓЃЈДгШШДјЕНББМЋКЃбѓЃЉЧГгк200mЫЎгђЩњЛюЕФ3916жжКЃбѓЧАШњбЧИйИЙзуЖЏЮяЮяжжЪ§ОнЃЌЗЂЯжЖЋЬЋЦНбѓКЭЮїДѓЮїбѓЕФЖрбљадМАЖрбљадЬнЖШЃЈЭМ18ЃЉЖМЗЧГЃЯрЫЦЃЌОЁЙмЖўИіКЃгђМфДцдкаэЖрживЊЕФЮяРэМАРњЪЗЕФВювьЁЃжЕЕУзЂвтЕФЪЧЃЌЮяжжзюЗсИЛЕФЧјгђВЂВЛдкГрЕРИННќЃЌЖјДѓдМдкЮГЖШ10-25ЖШжЎМфЁЃ

ЭМ18 ЖЋЬЋЦНбѓКЭЮїДѓЮїбѓКЃбѓЧАШњбЧИйИЙзуЖЏЮяЮГЖШЖрбљадЬнЖШЃЌЪ§ОнЮЊЕЅЮЛЮГЖШЕФЮяжжЪ§ЃЈв§здRoy et al. 1998ЃЉ
OberdorffЕШЃЈ1995ЃЉЪеМЏСЫШЋЧђЗЖЮЇЃЈЗЧжоЁЂУРжоЁЂбЧжоЁЂДѓбѓжоЕШЃЉЕФ292ЬѕКгСїЕФгуРрзЪСЯЃЌПчдНДгГрЕРЕНМЋЕиЕФЮГЖШЬнЖШЃЌЗЂЯжзюИпЕФЮяжжЖрбљадЮЛгкГрЕРИННќЕФЕЭЮГЖШЕиЧјЃЌЫцЮГЖШЩЯЩ§гуРржжРрЪ§УїЯдЯТНЕЕФЧїЪЦЃЌЫфШЛгаЯрЕБЕФБфЛЏЗљЖШЃЈЭМ19ЃЉЁЃ
ЭМ19 ЪРНчКгСїЕЫЎгуРрбиЮГЖШЕФЖрбљадЬнЖШЃЈЗТOberdorff et al.1995ЃЉ
2.ЮТЖШаЇгІЁЊЫцзХЫЎЮТЕФЩ§ИпЃЌЮяжжЖрбљадж№НЅдіМг
RoyЕШЃЈ1998ЃЉЗжЮіСЫЖЋЬЋЦНбѓКЭЮїДѓЮїбѓЧГЫЎЫЎгђЩњЛюЕФ3916жжКЃбѓИЙзуЖЏЮяЮяжжгыФъЦНОљКЃУцЮТЖШжЎМфЕФЙиЯЕЃЌЗЂЯжЖўепжЎМфДцдкУмЧаЕФЯрЙиЙиЯЕЃЌМДЕБКЃУцЮТЖШГЌЙ§20oCКѓЃЌЮяжжЪ§ДѓЗљдіМгЃЌЖјЧвЖўИіКЃгђЕФЧїЪЦОљЗЧГЃЯрЫЦЃЈЭМ20ЃЉЁЃ
ЭМ20 ФъЦНОљКЃУцЮТЖШКЭЮїДѓЮїбѓКЭЖЋЬЋЦНбѓКЃбѓИЙзуЖЏЮяЕФЮяжжЖрбљадЃЈвд1oЮГЖШЮЊМфИєЃЉЃЈв§здRoyet al. 1998ЃЉ
3.ЫЎЩюаЇгІЁЊНјШыЩюКЃЧјЃЌЮяжжЪ§МБОчЯТНЕ
ТНЕиЩЯКЃАЮИпЖШгАЯьТНЩњЖЏЮяЕФЮяжжЖрбљадИёОжЃЌдкКЃбѓЩњЬЌЯЕЭГжаЃЌЩюЖШЖдгуРрЕФЮяжжЗжВМИёОжвВВњЩњЯджјгАЯьЁЃдквЛаЉЧщПіЯТКЃбѓЩњЮяЕФЮяжжЖрбљадЫцЫЎЩюЕФдіМгЖјЯТНЕЃЌдкСэвЛаЉЧщПіЯТШДЪЧЯШЩЯЩ§КѓЯТНЕЁЃ
MacphersonКЭDuarteЃЈ1994ЃЉЪеМЏСЫДѓЮїбѓЖЋВП1746жжгуРрзЪСЯЃЌЗжЮіСЫЩюЖШКЭЮГЖШгыЕзЦмгуРрЮяжжЖрбљадЕФЙиЯЕЃЌЗЂЯжЕзЦмгуРрЕФЮяжжЪ§ЫцЮГЖШЕФдіМгЖјЯТНЕЃЈДѓдМЮГЖШУПЩЯЩ§5oЃЌгуРржжРрЪ§МѕЩй1%ЃЉЃЌЕЋЪЧдкДѓЖрЪ§ЮГЖШЃЌЮяжжЪ§дкДѓдМ150-300 mЩюЖШДяЕНзюДѓЃЈЭМ21ЃЉЁЃ
ЭМ21 ДѓЮїбѓЖЋВПЕзЦмгуРрЮяжжЖрбљадгыЮГЖШКЭЩюЖШжЎМфЕФШ§ЮЌЙиЯЕЃЈЗТMacpherson & Duarte 1994ЃЉ
SvavarssonЕШЃЈ1993ЃЉбаОПСЫХВЭўЁЂИёСжРМЁЂБљЕККЭББМЋКЃЕФКЃбѓЕШзуРрЃЌЙВга106жжЃЌжївЊЗжВМдкЧГЫЎКЃгђЃЌЫцзХЩюЖШЕФдіМгжжРрЪ§МБОчМѕЩйЃЈЭМ22ЩЯЃЉЁЃMorentaЕШЃЈ1998ЃЉДгBaleancЕКФЯВПЃЈЮЛгкЕижаКЃЮїВПЕФАЂЖћМАРћбЧЭхЃЉЫЎЩю200-1800mЧјгђВЩМЏСЫ82жжгуРрЃЌЗЂЯжжжРрЪ§ЫцЫЎЩюЕФдіМгЖјУїЯдМѕЩйЃЈЭМ22ЯТЃЉЁЃ
ЭМ22 ББВПКЃгђЕШзуРрЮяжжЖрбљадвдМАBaleancЕКДѓТНаБЦТгуРржжРрЪ§ЫцЫЎЩюЕФБфЛЏЃЈЗТSvavarsson et al. 1993, Morenta et al. 1998ЃЉ
4.РњЪЗЕФЕиРэИёОжЁЊвРШЛЪЧЮГЖШдНЕЭЃЌЮяжждНЗсИЛ
ЮяжжЖрбљадбиЮГЖШЕФЗжВМИёОжВЂВЛЪЧФЉДЮБљЦквдРДЖРгаЕФЯжЯѓЃЌЭМ23ЪЧДѓдМ7000ЭђФъЧАКЃбѓгаПЧГцРрЛЏЪЏЮяжжбиЮГЖШЬнЖШЕФЗжВМИёОжЃЌвВЪЧдкЕЭЮГЖШЕиЧјЮяжжЪ§зюЮЊЗсИЛЃЌЖјЯђИпЮГЖШЕиЧјЗНЯђЃЌЮяжжЪ§ШёМѕЁЃ
ЭМ23 дМ7000ЭђФъЧАЛЏЪЏКЃбѓгаПЧГцРрбиЮГЖШЬнЖШЕФЗжВМЃЈRosenzweig 1992аоИФздStehli et al. 1969ЃЉ
ЮхЁЂЮяжжЖрбљадЁЊвЛАуИёОжгыГЩвђ
1.ЮГЖШЁЂКЃАЮИпЖШКЭЪЊШѓЖШЁЊЫмдьСЫЮяжжЖрбљадЕФЛљБОИёОж
дкгАЯьЮяжжЖрбљадЕиРэИёОжЕФИїжжвђЫижаЃЌКСЮовЩЮЪвдЮГЖШЕФаЇгІзюЮЊЯджјЃЌвВзюЪмЙизЂЃЛКЃАЮИпЖШКЭЪЊШѓЖШвВЪмЕНЙуЗКЙизЂЁЃбизХетаЉЛЗОГвђзгЬнЖШЃЌЮяжжЖрбљЕиРэИёОжГЪЯжГівЛаЉЛљБОИёОжЃК
1ЃЉЮГЖШЪЧгАЯьЖЏЮяЃЈЫЎЩњЛђТНЩњЃЉКЭжВЮяЮяжжЖрбљадИёОжЕФживЊвђЫиЃКЮяжжЖрбљадЫцЮГЖШЕФЩЯЩ§ЖјЯТНЕЃЌвВМДЕЭЮГЖШЕФШШДјЕиЧјЕФЮяжжЖрбљаддЖИпгкЮТДјЕиЧјЃЌЖјЮТДјЕиЧјгждЖИпгкМЋЕиЁЃЕБШЛЃЌЩцМАЬиЖЈРрШКЪБПЩФмЛсгавЛаЉР§ЭтЃЌЦЉШчгаЕФРрШКдкжаЮГЖШЕиЧјзюДѓЃЌгааЉРрШКдђПЩФмжївЊОжЯогкМЋЖЫКЎРфЕФЦјКђЁЃвђДЫЃЌвЛаЉРрШКЕФЦ№дДЗжЛЏЁЂЩњРэЩњЬЌЪЪгІадЕШПЩОіЖЈЦфЬиЪтЕФЮяжжЗжВМЕиРэИёОжЁЃ
2ЃЉКЃАЮИпЖШЭЈЙ§ДЙжБЮТЖШБфЛЏгыЮГЖШЕФЕШаЇадЃЌЖдЮяжжЖрбљадвВВњЩњЯджјЕФгАЯьЃЌМДЮяжжЖрбљадвЛАуЫцКЃАЮЕФЩЯЩ§ЖјЯТНЕЁЃ
3ЃЉЪЊШѓЖШвВЗЧГЃЯджјЕигАЯьжВЮяЕФЮяжжЖрбљадЃЌЬиБ№ЪЧдкШШДјЁЊбЧШШДјЕиЧјЃЌИЩКЕЕиЧјЕФЮяжжЖрбљаддЖЕЭгкЪЊШѓЕиЧјЃЌШчИЩВндЕФЮяжжЖрбљаддЖЕЭгкШШДјгъСжЁЃ
4ЃЉЮГЖШЛђКЃАЮИпЖШЁЊЪЊЖШСЊКЯаЇгІЫмдьСЫМЋЖЫВЛЭЌЕФЬиеїаджВБЛРраЭЁЊЖЏжВЮяЮяжжМЋЮЊЗсИЛЕФШШДјгъСжЃЌЖЏжВЮяЮяжжМЋЮЊЦЖёЄЕФЖГдКЭЩГФЎЁЃ
2.ЮяжжЖрбљадИёОжЁЊЩњЬЌЁЂНјЛЏКЭРњЪЗЕФВњЮя
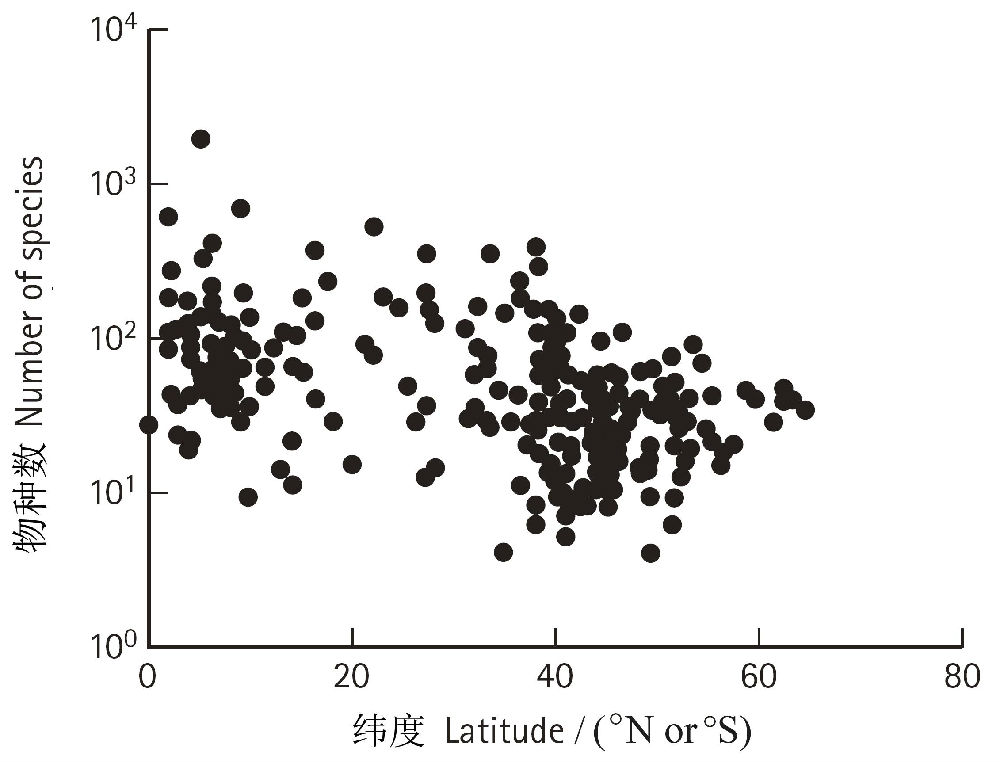
ЖдДѓЖрЪ§ЃЈТНЩњЛђЫЎЩњЃЉЖЏжВЮяРрШКЃЌЕЭЮГЖШЕиЧјЕФЮяжжЖрбљадБШИпЮГЖШЕиЧјвЊИпЕУЖрЃЌШчЙћАќКЌЕФРрШКдНЙуЗКЃЌетжжЧїЪЦдНУїЯдЁЃЮЊКЮШШДјЕиЧјБШЮТДјЕиЧјЩњЮяЖрбљадвЊИпЃПGastonКЭSpiceЃЈ2004ЃЉСаОйСЫШ§жжПЩФмЕФЛњжЦЃЌШЯЮЊетШ§жжЛњжЦПЩвдСЊКЯзїгУЃЌЫфШЛвВЛсЪмЕНвЛаЉЦфЫќвђЫиЕФгАЯьЃК
1ЃЉУцЛ§аЇгІЃЈAreaeffectsЃЉЃК RosenzweigЃЈ1992ЃЉШЯЮЊШШДјЧјгђЕФУцЛ§БШЮТДјДѓЃЈЖјЧвГрЕРИННќЕФШШДјЪЧНєУмЯрСЌЕФЃЉЃЌвђДЫЕМжТНЯИпЕФЗжЛЏЫйТЪКЭНЯЕЭЕФУ№ОјЫйТЪЁЃ
2ЃЉФмСППЩРћгУадЃЈEnergyavailabilityЃЉЃКЕЭЮГЖШЕиЧјгаИќЖрЕФФмСПЙЉИјЃЌвђДЫОпгаИќЗсИЛЕФзЪдДЃЌПЩвджЇГХИќЖрЕФЮяжжЁЃ
3ЃЉгааЇНјЛЏЪБМфЃЈeffectiveevolutionary timeЃЉЃКДѓЗЖЮЇЕФЛЗОГШХЖЏЃЈШчБљЦкЁЂЦјКђИЩКЕЕШЃЉЖдШШДјЕФгАЯьНЯаЁЃЌвђДЫЬсЙЉСЫИќЖрЕФгааЇНјЛЏЪБМфЃЈОјЖдНјЛЏЪБМф+НјЛЏЫйТЪЃЉЃЌДЫЭтЃЌЕижЪРњЪЗЪБЦкШШДјЕиЧјЕФУ№ОјЫйТЪвВПЩФмЯрЖдНЯЕЭЁЃ
ашвЊжИГіЕФЪЧЃЌШШДјЕиЧјЕФТНгђУцЛ§ЫфДѓгкЮТДјЕиЧјЃЌЕЋНіППетжжУцЛ§ЩЯгаЯоЕФВювьЃЈЭМ24ЃЉЪЧЮоЗЈШЋВПНтЪЭШШДјЕиЧјКЭЮТДјЕиЧјЮяжжЪ§ЕФОоДѓВювьЕФЃЌЛЙашвЊМгЩЯНЯЕЭЕФУ№ОјЫйТЪКЭНЯИпЕФЮяжжЗжЛЏЫйТЪЁЃЦфЪЕШШДјЕиЧјЗжЛЏЫйТЪНЯПьгІИУЪЧПЩвдРэНтЕФЃЌдкИпЮТЃЈЕБШЛЛЙгаЪЪвЫЕФЫЎЗжЙЉИјЃЉЬѕМўЯТЃЌЩњЮяЕФЩњГЄЗБжГЫйТЪНЯПьЃЌвХДЋБфвьКЭЮяжжЗжЛЏЕФЫйТЪвВгІИУИќПьЁЃДЫЭтЃЌШШДјЕиЧјИДдгЕФЮяжжМфЯрЛЅзїгУвВПЩФмЪЧЮяжжЗжЛЏЫйТЪНЯИпЕФдвђжЎвЛЁЃ
ЭМ24 ТНЕиЯрЖдУцЛ§КЭЮГЖШЕФЙиЯЕЃЈЗТRosenzweig 1992ЃЉ
MittelbachЕШЃЈ2007ЃЉЛљгкЩЯЪіШ§жжЛњжЦЃЌЬсГіСЫЯргІЕФМйЩшФЃаЭЃЈЭМ25ЃЉЃЌЕквЛжжМйЫЕШЯЮЊЃЌШШДјКЭЮТДјдкЮяжжЗжЛЏЫйТЪЩЯВЂЮоВювьЃЌЕЋгЩгкЩњЬЌвђЫиЃЈШчЩњЬЌЮЛЃЉЕФВювьЃЌШШДјЖдЮяжжЕФДцдиСІИпгкЮТДјЃЛЕкЖўжжМйЫЕШЯЮЊЃЌШШДјЕФЮяжжЖрбљЛЏЫйТЪЃЈЗжЛЏЫйТЪЁЊУ№ОјЫйТЪЃЉБШЮТДјИпЃЌвђДЫЮяжжЛ§РлЫйЖШНЯПьЃЛЕкШ§жжМйЫЕШЯЮЊЃЌШШДјБШЮТДјОпгаИќЖрЕФЪБМфНјааЮяжжЖрбљЛЏЃЌЦЉШчЃЌЭМжаМ§ЭЗБэЪОИпЮТЦјКђЕФНсЪјКЭЮТДјЕФаЮГЩЁЃ

ЭМ25 НтЪЭЮяжжЖрбљадЮГЖШЬнЖШЕФШ§жжМйЫЕЃКЃЈaЃЉЩњЬЌМйЩшЃЌОлНЙЮяжжЙВДцКЭЮяжжЖрбљадЕФЮЌГжЛњжЦЃЌЃЈbЃЉНјЛЏМйЫЕЃЌОлНЙЮяжжЗжЛЏЫйТЪЃЌЃЈcЃЉРњЪЗМйЫЕЃЌЕиЧђРњЪЗжаШШДјЛЗОГЕФГжајЪБМфКЭЗЖЮЇЃЈЗТMittelbach et al. 2007ЃЉ
Нсгя
ЮяжжЪЧЩњУќЩњДцЁЂЗБбмгыНјЛЏЕФЛљБОЕЅдЊЃЌЕиЧђЩЯвбУшЪіЕФЮяжжГЌЙ§СЫ170ЭђЁЃШчДЫЗБЖрЕФЮяжжЪЧдѕбљБЛДДдьГіРДЕФЃПЮоТлЪЧРТэПЫгУНјЗЯЭЫКЭЛёЕУадвХДЋЃЌЛЙЪЧДяЖћЮФЕФЫцЛњБфвьЁЂЩњДцОКељКЭздШЛбЁдёЃЌЛђЪЧЛљгкЛљвђЭЛБфЕФЭЛБфТлЃЌЖМжЛЪЧЙизЂЮяжжДДдьЕФЮЂЙлЙ§ГЬЃЈКСЮовЩЮЪетЪЧживЊЕФЮЂЙлЛљДЁЃЉЁЃЕЋЪЧЃЌДгКъЙлЕФЪгНЧРДПДЃЌЮяжжИќЪЧЕигђадЩњЬЌЛЗОГРњЪЗЙ§ГЬЕФВњЮяЃЌвђДЫВХаЮГЩСЫЮяжжЖрбљадЕФЕиРэИёОжЁЃ
МђбджЎЃЌЮяжжОЭЪЧгъЁЂШШЕФВњЮяЃЌЕиЧђЩЯгъЁЂШШЕФЗжВМИёОжОЭОіЖЈСЫЮяжжЖрбљадЕФКъЙлИёОжЃЌМДзюЮЊЪЊШШЕФЕиЧјЃЈШчШШДјгъСжЃЉЩњВњСІзюИпЃЌЮяжжЗжЛЏзюПьЃЌвђДЫжЇГХЕФЮяжжзюЮЊЗсИЛЖрбљЁЃЮяжжЖрбљадЕФЕиРэИёОжухдЁЙиМќЛЗОГвЊЫиЃЈЮГЖШЁЂКЃАЮИпЖШКЭЪЊШѓЖШЃЉЕФВЛЖЯЫмдьЃЌДгБОжЪЩЯПДЪЧЩњЬЌЁЂНјЛЏКЭРњЪЗЙ§ГЬЕФВњЮяЁЃ
етбљЃЌШЋЧђЦјКђФЃЪНЭЈЙ§КъЙлЕФЩњЬЌЙ§ГЬЃЈЕБШЛБиаыдкЮЂЙлЕФГЩжжЛњжЦВйПиЯТЃЉМШОіЖЈСЫжВБЛЕФЕиРэЗжВМгыбнЬцФЃЪНЃЌвВжїдзСЫЮяжжЖрбљадЕФЕиРэИёОжЁЃетвЛЗНУцебЪОСЫЮяжжбнЛЏдкКъЙлЩЯЕФЗНЯђадЃЌЕЋВЂВЛЗёШЯЮЂЙлГЩжжЕФЫцЛњадЃЌСэвЛЗНУцвВИцЫпЮвУЧВЛЭЌЕижЪФъДњгЩгкЦјКђФЃЪНЕФОоБфЛсГіЯжЭъШЋВЛЭЌЕФЩњЮяЧјЯЕЃЌетЭљЭљПЩФмЛсИјЙХЩњЮябЇМвСєЯТЕигђадЛЏЪЏМЧТМВЛСЌајадЕФБэЯѓЃЌВЂБЛЦфгУРДЕБзїЬјдОНјЛЏЕФЁАПЩППЁБжЄОнЁЃ
жївЊРДдДЃК
аЛЦН. 2013. ДгЩњЬЌбЇЭИЪгЩњУќЯЕЭГЕФЩшМЦЁЂдЫзїгыбнЛЏЁЊЩњЬЌЁЂвХДЋКЭНјЛЏЭЈЙ§ЩњжГЕФШкКЯ. ББОЉЃКПЦбЇГіАцЩч
ЃЈгЂЮФЃКXie P. 2013. ScalingEcology to Understand Natural Design of Life Systems and Their Operations andEvolutions ЈC Integration of Ecology, Genetics and Evolution through Reproduction. Beijing: Science PressЃЉ
в§ЪіИУВЉЮФЕФЯрЙиФкШнЪБЃЌЧыв§гУИУзЈжјЁЃ
ЕчзгАцЯТдиЃКhttp://wetland.ihb.cas.cn/lwycbw/qt/
ЯрЙиВЉЮФЃК
ЮЊКЮФкУЩЙХХћЙќЮолѓВндЃЌЖјбЧТэбЗгъСжНПцЌЃП
http://blog.sciencenet.cn/home.php?mod=space&uid=1475614&do=blog&id=815185
НтЖСЩњУќЕФЩшМЦЁЊЮЊКЮЫќЮЂШчГОАЃврОожСДѓЯѓЃП
http://blog.sciencenet.cn/blog-1475614-808322.html
ДяЖћЮФНјЛЏТлжЎбяЦњЃЌЩњУќЦ№дДРэТлжЎИяаТЃЁ
http://blog.sciencenet.cn/blog-1475614-805836.html
https://wap.sciencenet.cn/blog-1475614-816369.html
ЩЯвЛЦЊЃКЮЊКЮШЫЁАОхХТЁБНќНЛЖјжВЮяШДЪШКУЃП
ЯТвЛЦЊЃКжВЮяЁАадЁБЕФбнНјЁЊЁЊКљТЋРяТєЕФЪВУДвЉЃП
75 ГТСС ЗћЛд ЭѕЕТЛЊ КТРж ПТжОаТ ЭѕКЭдЦ ФпРжвт ЩђКъ ГТПЌКВ РюЧсжл бюе§ъВ еХЯіСж едЕЄЕЄ жьЙЂЦН еХФмСЂ Н№Ие амйЛ дЌОќЗЈ ЛЦгРвх РюЭў зЃЙњШй РюЭСШй ЭѕШйСж аЄРя ВЬбу Сѕбѓ ЕЫЕРЙѓ РюаЁЮФ дЌЙ№Яу еХДѓЮФ ГТОДЦг ЮтвЋ КњОХСњ ёћНЃъЭ БЋЕУКЃ еХбхБѓ ГЬЦ№ШК еХцї И№ЮЌбЧ еХдЦбя гІааШЪ едБѓ СКИпЕР СѕЙтвј СѕБјЧе ЖНрСж ЮтЪРП ЪЏХэСщ ЙњЯўДК РюРђ ЭѕЛЖ еХУШ И№Зх ЧПЬЮ РюгБвЕ грЕУеб wangqinling eastHL2008 ddsers yyzhao1104 wanghua4 rfm2007 scr3322 yanghua7788 tangxin aliala farniu leihh Lazi711 pangxinghong88 liguangyu2001 sgcst fishstop0910 sjtzcqn capenter
ИУВЉЮФдЪаэЪЕУћгУЛЇЦРТл ЦРТл (32 ИіЦРТл)
ШЋВПзїепЕФОЋбЁВЉЮФ
- • ЮЊБЃЛЄЩњЮяЕиРэбЇПЊЦєвЛЩШаТДАЛЇ
- • вЛИіЛљгкКъбнЛЏжЄОнЕФецКЫЩњЮяЦ№дДаТРэТлЁЊЁЊDNAбЙЫѕМйЫЕЃЈThe DNA Packing HypothesisЃЉ
- • ЗЧОЕфЩњЮяВйзнЃКИЛгЊбјЕЫЎЫЎЬхжаСІПЫРЖдхЫЎЛЊЕФЧПСІЩњЬЌММЪѕ
- • дкДѓАгжмБпПЊБйНќздШЛЕФХдЭЈЕРНЋЮЊіргуЕФБЃЛЄЕуССЯЃЭћ
- • ЁАФЇЗЈЬиеїЁБаТР§жЄЁЊЁЊЖЋбЧЬигаР№ПЦгуРрЪЪгІЗјЩфжаВњТбЯАадбнЛЏЛњжЦ
- • дЫгУЗЧОЕфЩњЮяВйзнЗРПиЮфККЖЋКўРЖдхЫЎЛЊЕФГЩЙІЪЕМљ
ШЋВПзїепЕФЦфЫћзюаТВЉЮФ
- • ЗЧОЕфЩњЮяВйзнРэТлМАЦфГЩЙІгІгУШыбЁ2023ФъЖШКўББЪЎДѓПЦММНјеЙ
- • ЮЊБЃЛЄЩњЮяЕиРэбЇПЊЦєвЛЩШаТДАЛЇ
- • вЛИіЛљгкКъбнЛЏжЄОнЕФецКЫЩњЮяЦ№дДаТРэТлЁЊЁЊDNAбЙЫѕМйЫЕЃЈThe DNA Packing HypothesisЃЉ
- • ЗЧОЕфЩњЮяВйзнЃКИЛгЊбјЕЫЎЫЎЬхжаСІПЫРЖдхЫЎЛЊЕФЧПСІЩњЬЌММЪѕ
- • ЁАПеЦјИЛгЊбјЛЏЁБИјШЫРрНЁПЕДјРДаТЭўаВ
- • дкДѓАгжмБпПЊБйНќздШЛЕФХдЭЈЕРНЋЮЊіргуЕФБЃЛЄЕуССЯЃЭћ
ШЋВПОЋбЁВЉЮФЕМЖС
- • Ь§ЕМЪІЕФЛАЃЌБЯвЕТлЮФД№БчетбљзМБИ
- • ПЦбЇМвУЧРЇЛѓСЫ50ЖрФъЃЌвЛИіЪЅБАуЕФЛЏбЇжЎУежегкНтПЊСЫ
- • ЕТЙњTU IlmenauРзгТЕШзлЪіЃКгУзїЯШНјЕчЛЏбЇДЂФмВФСЯЕФЮЂФЩжЇГХНсЙЙЁЊзюаТНјеЙЁЂбаОПЬєеНКЭЮДРДЧАОА
- • ЩѓИхШЫЧПжЦзїепв§гУздМКЭХЖгЮФЯзЕФЬзТЗ
- • ЮвКЭЧЎРэШКЕФвЛУц
- • УћМвВЉЪПТлЮФ36ЃКИпЗжзгЛЏбЇМвЛЦнсЭЌдКЪПВЉЪПТлЮФ1953ФъВМТГПЫСжРэЙЄбЇдК
ЯрЙиВЉЮФ
- • УРЙњзєжЮбЧДѓбЇЕШЛњЙЙбЇепЃКизИюВпТдЖдBulldog 805зЯЛЈмйоЃ+Tifton 85ЙЗбРИљЛьВЅВнЕиВњСПМАЦЗжЪЕФгАЯь
- • УРЙњПАШјЫЙжнСЂДѓбЇЁЂУмЫеРяДѓбЇЕШЛњЙЙбЇепбаОПГЩЙћЃКЭСШРЫЎЗжЙмРэВпТдКЭЦЗжжЖрбљадЖдзЯЛЈмйоЃВњСПЁЂгЊбјЦЗжЪКЭХЉГЁгЏРћФмСІЕФгА
- • ЕТЙњЁЂНнПЫВнвЕПЦбЇбЇепГЄЦкЗХФСЪЕбщЃКвьжЪВнЕиАпПщжаЕФЭСШРгаЛњЬМДЂСПКЭЕиЯТЩњЮяСП
- • ШчКЮВХФмСЌајБЛЦРЮЊЪЎФъЁАжаЙњИпБЛв§бЇепЁБЃПЁЊЁЊЖдЛАЩЯКЃДѓбЇЭѕЧфЮФНЬЪк
- • C2CЃКФЯВ§ДѓбЇбЇепЬсГібЇЪѕНЛСїаТИХФю
- • бЇепЮЊЪВУДЕУЗЂБэЮФеТЃП