博文
自然选择:谁选择?为何选择?选择什么?
 精选
精选
|
美国古生物学家和进化生物学家Stephen Jay Gould(1941-2002)在“Ever Since Darwin: Reflections in Natural History”(中译本:达尔文以来—自然史沉思录)中说,“为什么自然选择被杜布赞斯基比作作曲家,被辛普森比作诗人,被迈尔比作雕刻师,以及被朱安利·赫胥黎比作芸芸众生的莎士比亚?我不想为这些比喻的挑选做辩护。但是我愿支持这种倾向,即说明达尔文主义的本质就是自然选择具有创造性”(Gould 1977)。巴顿等(2010)宣称,“自然选择是唯一导致适应的过程……是一种重要的进化过程”。但是,对达尔文的自然选择也有不少的疑虑。譬如,Robson和Richards(1936)曾说,“我们不相信,在进化中作为一个可能的因素的自然选择是可以忽视的。然而,却很少正面的证据对它是有利的……以致使我们没有权力来确定它在进化中有主要的创造性作用”。
1. 自然选择概念的历史回顾
达尔文指出,古代学者对自然选择只是影射性地述及过:亚里士多德在《听诊术》(Physic Auscultationes)中说,“有什么会阻止身体的不同部分去发生自然界中这种偶然的关系呢?例如,牙齿为了需要而生长了,门齿锐利,适于分切食物,臼齿平钝,适于咀嚼食物,它们不是为了这等作用而形成的,这不过是偶然的结果而已。身体的其他部分亦复如此,它们的存在似乎是适应一定目的的。因此,所有一切构造(即一个个体的所有部分)都好像是为了某种目的而被形成的,这一切经过内在的自发力量而适当组合之后,就被保存下来了,凡不是如此组合而成的,就灭亡了,或趋于灭亡”,从这里可看到自然选择原理的萌芽,但亚里士多德对这一原理还没有充分的了解(Darwin 1872)。
法国唯物主义哲学家狄德罗(Denis Diderot ,1713 —1784年)在其1749年出版的《盲人通讯》中曾说,在地球历史的开始,自然试着通过自然发生产生出许多生命类型,其中许多类型肯定缺少基本的生命器官,所有这些畸形都会死掉,但是偶尔会有机遇降临到一个能够生活下去和生殖的类型上,因此,有些物种之所以能够生活在地球上,是经过了一个试错的过程(鲍勒1999)。鲍勒认为,狄德罗假设的是淘汰自然发生中的粗陋物质而不是已经确立的物种的前进过程,因此,与自然选择学说不太一样。
据达尔文记载,Wells H C于1813年在皇家学会宣读了一篇题为《一位白种妇女的局部皮肤类似一个黑人皮肤的报告》的论文,其中说到,一切动物在某种程度上都有变异的倾向,而农学家们利用选择来改进他们的家养动物,“人工选择所曾完成的,自然也可以同样有效地做到,以形成人类的一些变种,适应它们所居住的地方,只不过自然选择比人工选择来的徐缓而已。最初散居在非洲中部的少数居民,可能发生一些偶然的人类变种,其中有的人比其他人更适于抗拒当地的疾病。结果,这个种族的繁衍增多,而其他种族则将衰减;这不仅由于他们无力抗拒疾病的打击,同时也由于他们无力同较为强壮的邻族进行竞争。如上所述,我认为这个强壮种族的肤色当然是黑的。但是,形成这些变种的同一倾向依然存在,于是随着时间的推移,一个越来越黑的种族就出现了:既然最黑的种族最能适应当地的气候,那么最黑的种族在其发源地,即使不是唯一的种族,最终也会变成占优势的种族”,然后他又把同样的观点引伸到居住在气候较冷的白种人(Darwin 1872)。达尔文承认这是对自然选择原理的最早认识,但认为仅局限于人种的某些性状。依笔者之见,Wells其实已将达尔文进化论的核心思想(随机变异、生存斗争和自然选择,其结果是适者生存)一网打尽!
植物学家诺丁(Naudin M)在一篇于1852年发表的讨论物种起源的论文中认为,物种形成的方式同变种在栽培状况下形成的方式是类似的,他把后一形成过程归因于人类的选择力量,但他没有阐明选择在自然状况下是怎样发生作用的。他说,这是“一种神秘的、无法确定的力量,对某些生物而言,它是宿命的;对另外一些生物而言,它乃是上帝的意志。为了所属类族的命运,这一力量对生物所进行的持续作用,便在世界存在的全部时期内决定了各个生物的形态、大小和寿命,正是这一力量促成了个体和整体的和谐,使其适应于它在整个自然机构中所担负的功能,这就是它之所以存在的原因”(Darwin 1872)。
生命个体既选择也被选择,前者是常识,而后者却不那么直观。即便如此,被选择的朴素思想也是源远流长的。达尔文的追随者杜布赞斯基(1964)客观地评价道:“曾经在古老的经典中预示过的自然选择(naturalselection)原理,由达尔文把它提高到了科学理论的地位上”。
2.谁选择?为何选择?
达尔文深信人工选择的威力,这个选择者就是饲养家,他说,“饲养者习惯说动物的体制好像是可塑性的东西,几乎可以随意塑造”,并引用了动物鉴定家尤亚特(Youatt)的话:人工选择“可以使农学家不仅能够改变他的畜群性状,而且能够使它们发生完全的变化。选择是魔术家的魔杖,用这只魔杖,可以随心所欲地把生物塑造成任何类型和模式”(Darwin 1872)。达尔文认为,自然选择所不能做的是:改变一个物种的构造,而不给它一点利益,却是为了另一个物种的利益。他以几种不同的方式来述说他的自然选择:
①“我把每一个有用的微小变异被保存下来的这一原理称为‘自然选择’,以表明它和人工选择的关系。但是,斯宾塞先生所常用的措词‘最适者生存’,更为确切,并且有时也同样方便……”自然选择“是一种不断活动的力量,它无比地优越于微弱的人力,其差别正如‘自然’的工作和‘人工’相比一样”;
②“每一物种所产生的个体,远远超过其可能生存的个体,因而便反复引起生存斗争,于是任何生物所发生的变异,无论多么微小,只要在复杂而时常变化的生活条件下以任何方式有利于自身,就会有较好的生存机会,这样便被自然选择了。根据强有力的遗传原理,任何被选择下来的变种都会有繁殖其变异了的新类型的倾向”。
③“我把这种有利的个体差异和变异的保存,以及那些有害变异的毁灭,叫作‘自然选择’,或‘最适者生存’。无用也无害的变异则不受自然选择的作用,它或者成为彷徨的性状,有如我们在某些多形的物种里所看到的,或者终于成为固定的性状,这是由生物的本性和外界条件来决定的”。
迈尔(2009)如此解读道:“谁来选择?在人工选择中,是动植物培育者挑选出一些优秀的个体作为下一代的种源。但是,严格地说,在自然选择中没有这样的代理者。实际上达尔文将自然选择称作一个淘汰的过程。作为下一代亲本的个体是那些由于具备或缺少某些性状因此比起同代的其他个体更能适应当时环境条件的个体。它们的兄弟姐妹都在自然选择过程中遭到了淘汰。赫伯特·斯宾塞曾经说过,自然选择只不过是‘适者生存’,他这样说是对的。自然选择是一个淘汰过程,达尔文在他的著作中采纳了斯宾塞的比喻”。
也有学者从遗传的观点出发,将自然选择定义为变异(或突变)类型的差别增殖(Muller 1960)。换句话说,自然选择发生在基因型之间的适合度存在差异的时候。这里适合度这个名词代表生物体留下后裔的能力。对每一个基因型来说,适合度是以每一个个体的达到成熟的后裔数目来度量的(Kimura 1983)。道金斯(2005)认为,“自然选择是相互竞争的DNA将自己成功地垂直传递到物种档案的这样一种过程。‘DNA竞争对手’是指物种染色体地址中的其他不同内容。这些基因要比竞争对手更容易、更成功地保存于档案”。Falconer & Mackay(2000)指出,“亲本之间不同的基因型可能有不同的生殖力,如果是这样,则它们对形成下一世代的配子贡献也不会相等。通过这种方式,基因频率在传递过程中可能会改变。而且,新形成的合子之间的基因型可能有不同的存活率,因此到个体成熟且它们本身成为亲本的时候,新的世代中基因频率也可能会改变。这些过程称为选择”。
自然选择也被区分为正选择和负选择(Kimura 1983):“如果一个基因的突变型(简称突变体)在群体中产生,而且它增强携带它的个体的平均适合度,从而帮助它本身在群体中的散布,我们说这是正选择在工作。这是达尔文所假设的为生物界中适应进化的主要原因的那种自然选择型式,可以公正地称之为达尔文选择。被选择的基因并不必须是一个新的基因,它可以是以前的一个有害基因,由于环境的变化而成为有利的基因。在另一方面,如果突变对生存力或生殖力的有害效应而减少个体的适合度,那么它将易于从群体中淘汰掉。这种类型的选择可以称为负选择”。依笔者之见,这种区分既无新意,也无意义,因为如果没有“害”,何来“利”?反之亦然!
人工选择的威力是强大的,农作物的选育满足了人类对粮食的需求(图1A),对动物的选育则推动了畜牧业的发展。人类对动物行为的选择效果也是惊人的,譬如,有学者通过实验对果蝇的向光飞行能力进行了筛选,起初,果蝇只能以50px/s的速率飞行,将那些最强的4.5%的果蝇选择出来继续实验,经过100代后,飞行的平均速度提高了85倍,达到了4250px/s(图1B)。对狗的选育效果亦令人惊愕(图2)。
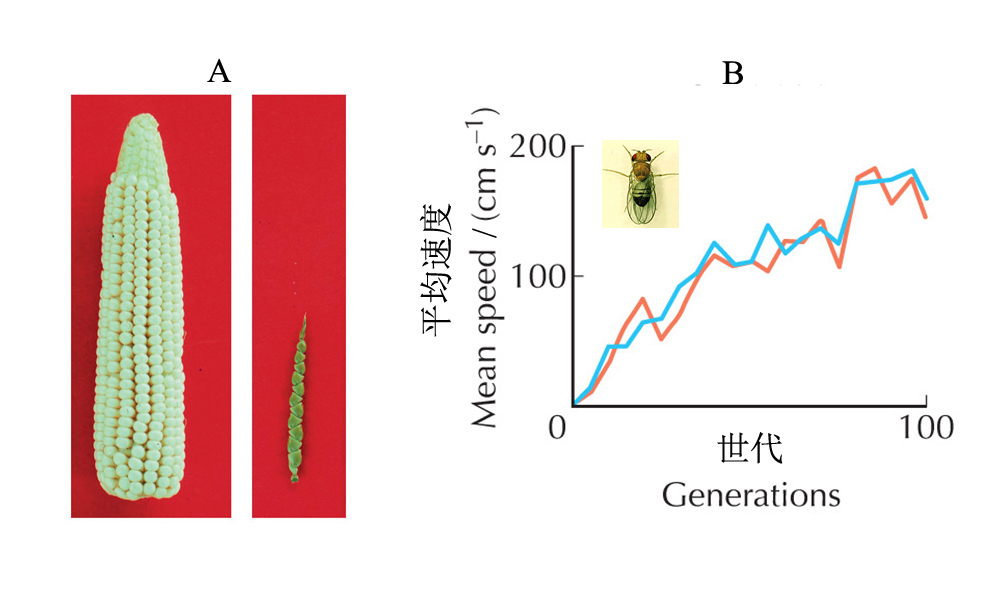
图1(A)农作物—玉米和与其有亲缘关系的野生品系—墨西哥类蜀黍的比较,(B)对果蝇飞行能力的选择,显示两个研究组的结果(引自巴顿等2010)

图2 奇瓦瓦和大丹:皮囊之下都是狼,但谁又能猜到经过几个世纪的人工选育后形态上发生了如此之大的变化(引自:Dawkins 2009)?
我的问题是,在没有人干预的自然界,到底谁进行了选择呢?或者说这个自然到底是什么呢?达尔文十分强调被选择者及其过程,但对选择者—自然及其动因并未耐心说明,到底自然为何要进行这样的选择或者说其目的何在呢?
道金斯(2005)指出,“自然选择—这个达尔文发现的盲目的、无意识的、自动的过程,我们今天所知的、对于生命的存在以及看似有意图的生命形式的解释—本身是毫无目的性的。它没有心智,也没有心智之眼,更不会对未来进行规划;它没有洞察力,没有先见之明,甚至根本没有‘视力’。如果非要说它在自然界中扮演了‘钟表匠’角色的话,那么它就是一个‘盲眼钟表匠’”。
巴顿等(2010)认为,自然选择创造了复杂的、更为适合的生命结构特征。但笔者认为未必都会如此,因为既有复杂化的情形,也有简化的情形(如寄生),适合也不是绝对的或永续的。正如杜布赞斯基(1964)所说,“自然选择只对在一个时候有利的变异体起作用,而不顾它们以后的价值如何。选择是没有预见性的……”。
3. 选择什么?
(1)基因选择
丹麦遗传学家约翰森(Wilhelm Johannsen,1857–1927)于1911年提出的两个重要概念——基因型(genotype)和表现型(phenotype),前者指一个生物体内的DNA所包含的所有基因的总和,而后者指一个生物个体在特定环境中呈现出的性状(形态结构、生理生化、行为习性,等等)的总和,或者说它是个体从受精卵向成体发育过程中基因组对环境不断响应而产生出的一系列表型的总合。基因是内在或潜在的,而表型是外在或现实的。一般认为,表现型是基因的产物,但笔者认为,在一定程度上,反之亦然。威廉斯(2001)指出,“在最终意义上,自然选择设计一个控制论意义上的抽象概念—基因,和一个统计学意义上的抽象概念—平均表现型适合度”。
环境对表现型的影响有时极为深刻,诱导有机体启动不同类型的形态发育。威廉斯(2001)列举了一些著名的例子:“有时一个较大的表现型差异也许是来自于不起眼的、转瞬即逝的生态学因素,更多地是通过体质环境中的一个关键变化的触发。在发育早期微不足道的食物改变,也许会使一个蜜蜂变成一个工蜂而不是一个蜂王。遗传信息的含义在一个工蜂的体内与在一个蜂王的体内是不一样的。在有尾两栖类中,变态现象的存在或缺乏相似地也是由一个食物的阈值决定的。海洋动物幼虫阶段的固着和变态与从合适的固着位点发出的刺激有关。植物受到某种日夜周期的影响,就会决定它在以后一段时期的开花情况”。
环境对表现型产生显著影响的另一个例子就是水毛茛(Ranunculus aquatilis),其茎叶在水下生长时呈细条状,而在水面上生长时则呈宽阔的裂片状(图3)。如此巨大的形态可塑性,真是令人惊叹!

图3 水毛茛(Ranunculus aquatilis)
一些学者认为自然选择的基本单位应该是基因而不是个体,因为遗传信息通过基因世代相传,因此进化最应该从基因的视角来考虑,称之为基因选择(gene selection)或自私基因(selfish gene)理论。这与传统的个体选择理论相对立。分子生物学中心法则(central dogma of molecular biology)被认为是基因选择理论的基石。基因选择理论的倡导者包括英国进化生物学家William Donald Hamilton(1936-2000)、英国生物学家Colin Pittendrigh(1918-1996)、美国进化生物学家George Christopher Williams(1926-2010),而1976年英国进化生物学家道金斯(Richard Dawkins, 1941—)的“The Selfish Gene(译文:自私的基因)”的登场使之达到了巅峰。
道金斯宣称,自然选择的不是物种、群体,甚至也不是个体,而是基因,他认为,生命,不仅仅是生物体,而是基因(DNA的简单片段),因为它们是稳定的并且可以复制的,所以就能生存下来,“基因犹如钻石,是永恒的。个体和种群就像天空中的浮云,或者沙漠中的尘暴”(Mills 2010)。威廉斯(2001)说,“体质的演替和所选择的生态龛是为基因服务的工具和战术”,但笔者认为,这样的说法未必有理,事实上它们都应该是服务于种族延绵的工具和战术!

R. Dawkins
基因选择还被细分为:对有害突变的纯化选择(purifying selection)、维持变异的稳定选择(balancing selection)以及能固定有利突变的定向选择(directionalselection)(巴顿等2010)。
如果性状与基因之间存在唯一的对应关系,基因选择理论或许成立,而遗憾的是,这并不是所有的情形,因为很多基因之间还存在相互作用。其一,存在所谓基因多效性(pleiotropy,即一个基因影响多个蛋白的表达)和多基因效应(polygene,即由多个基因控制某个表现型)等;其二,一个基因在某个特定的基因型中可能是有利的,但在另外的基因型中则可能是有害的。因此,选择绝不可能都是针对单个基因。也许可以想象针对基因群,就像个体水平的群选择一样。
巴顿等(2010)强调,“自然选择需要影响适合度的遗传变异,也就是说必须有可遗传的变异影响到生物体的生存和繁殖”。杜布赞斯基(1964)指出,“在某一环境下为不利的突变,可能在改变了的环境下变成为有利的。因为自然选择不是对于个别突变发生作用的,而是对于基因组合或基因型发生作用的;因此一个与某些基因一起存在时降低生活力的突变,当它被放置在一个不同的基因背景上时,可能就会增进生活力……假如一个群体是由若干遗传上有区别的类型所组成,则其中有些类型或许会比另一些类型产生较多的生存后裔。于是,在以后的世代中,某些基因、基因复合体和染色体结构,就将增多;而另外的一些,则将减少”。
威廉斯(2001)感叹道,“需要引起生物学家注意的并不是突变发生的频繁性,恰恰相反,倒是它们的稀罕性。我们还能在自然界的什么地方,发现这样一种复杂的、具有生物化学功能的分子,它们能够经历数百年或数千年的时间,在陆地温度中、在水分子的包围之下,能够生存下来,并且保持不变?它们不仅生存下来,而且还以极高的精确度复制自身。确实,这些过程只能通过这一假设才可以解释,那就是一个长期的、无情的选择是有利于稳定维持的……”。
(2)个体选择
一般来说,传统的博物学家或生物学家信奉个体选择(individual selection)理论,认为自然选择作用于个体水平,因此适应是为了有利于个体,从而导致个体间的选择。达尔文的进化理论就是构筑在个体水平的自然选择之上,他曾说,“同种的个体之间所进行的斗争几乎必然是最剧烈的,因为它们居住在同一区域内,需要同样的食物,并且还遭遇同样的危险”(Darwin 1872)。美国进化生物学家迈尔(Ernst Walter Mayr,1904-2005)认为,基因只是表现型的组成部分,而个体总体上的表现型(建立在基因型基础上)才是选择作用的实际靶子(迈尔2009)。
迈尔强烈反对基因选择理论,认为基因永远不会是选择的焦点,因为基因是由个体携带的,而个体的性状则来自成千上万个基因以及这些基因的无数的相互作用,因此,基因从来都不是单独行动的,所以选择永远不可能作用在一个基因上,或者说自然影响的不是一个单独的基因而是个体整个基因组的集体行动和相互影响,因此,不管一个基因有多麽适应,只要它所寄居的生物体不适应,这个基因便会消失(Mills 2010)。迈尔(2009)认为,“暴露给自然选择的是表现型,而基因并不直接暴露给自然选择……正是由于选择的靶子是表现型而不是基因型,基因库中才得以存在大量的遗传变异”。
(3)群体选择
个体间(种内或种间)的生存斗争是达尔文进化论的灵魂。在很多情形下,同种个体之间确实存在残酷的生存竞争,但也有不少物种的个体之间也能演化出良好的协助关系(图4)。如果通过少数个体的牺牲可以改善群体适应的话,将会有利于物种更长的延续,这便引伸出所谓“群体选择”的问题。这也被认为是对利它主义进化的一种解释。
图4在南非Kalahari沙漠上的一只猫鼬正在观察捕食者,如果看见捕食者正在接近,它将发出警告的叫声,通知群体中的其它同伴能够逃跑与隐藏,随后自己跟随它们(引自Allman 2009)
其实,群体选择的思想可以追溯到达尔文,因为他曾指出,“在社会性的动物里,自然选择能使各个体的构造适应整体的利益;如果这种被选择出来的变异有利于整体”(Darwin 1872)。
美国哲学家和史学家费斯克(John Fiske, 1842-1901)在1874年出版的《宇宙哲学大纲》中倡导了一种协作观点,即那些彼此合作的动物通常能够更成功地生存下去,于是出于本能的合作就得到了强化。他提出利他主义是人类进化的突出特征,在他看来,利他主义,即愿意为了他人而牺牲自己的行为,是物种最成功的进化策略,从而从根本上激发了人类文明的发展(鲍勒1999)。

J. Fiske
俄国地理学家克鲁泡特金(Pyotr Alexeyevich Kropotkin,1842-1921)在1902年出版的《互助,进化的一个要素》一书。他根据自己对野外生物的观察认为,自然界中存在的斗争迹象并不多,动物一般通过与其他动物的合作来改善生存的机遇,因此有理由宣称进化实际上通过增加每一代合作的水平而进行的,动物群中成功发展出来的性状可以遗传给后代,并在后代中得到强化。他认为互助是包括人类在内的一切物种得以保存下来并不断进化的主要因素,并称这一理论是对达尔文生存斗争学说的一种补充,为达尔文主义弥补了一个重大的空白。

P. A. Kropotkin
据杜布赞斯基(1964)记载,在20世纪40年代,一些学者认为合作在适应上的重要性也存在于有些并不形成高度组织的社会生物中,一个孤立的完全独立的个体,可以说是捏造的;事实上,大多数或者甚至所有的生物,那是生存在或多或少联合的团体中,维持这些联合的能力,就会造成一些合作,至少是“原始的合作”。20世纪60年代,英国动物学家VeroCopner Wynne-Edwards(1906-1997)正式提出了群体选择(group selection)的进化理论,认为自然选择作用于群体而不是传统的个体水平,这可能适合于一些形成社会结构的物种(Wynne-Edwards 1962)。其实,群体选择也得以个体选择为基础。

V. C. Wynne-Edwards
许多生物学家已经暗示,并且有相当数量的生物学家明确支持,由相互作用的个体所组成的群体也许以这样一种方式适应性地组织起来,即个体利益通过一种功能上的从属关系服务于群体利益,这种与群体有关的适应必定可归功于群体的自然选择(威廉斯2001)。群体选择可细分为两种类型:软性群选择(指在疏散的类群中选择)和硬性群选择(指在有密切社会关系的类群中选择),硬性群中的成员通过相互合作(通报敌情、分享食源以及联合抗敌等等)增加类群生存的机会。渔猎阶段的人类可能就是采用的这种模式(迈尔2009)。
杜布赞斯基(1964)指出:“在社会生物里,一个个体的生存率和生活力,只有当它对于该群落(Colony)或类群(Groups)的维持能发生作用时,则它在适应上才是重要的。这个事实在昆虫社会中是非常明显的,在其中,大多数的个体是完全不育的。在群体的成员之间,竞争完全消除,而由合作和互助所代替”。
Allee等(1949)认为,一个群体不单是一个集群的意义,而是一个具有空间和时间性的实体,它具有以下五种有机属性:(1)具有一定的结构和组成;(2)群体有其自己的个体发育,它象生物一样,表现着生长、分化和分工、生存、衰老以及死亡;(3)群体也具有遗传性;(4)群体是由相互依存的遗传和生态两方面的因素的作用下发展起来的;(5)群体象一个有机体一样,是作为一个整体单位而接受环境的影响的;这是一个相互的现象,由于这种影响的结果,引起了群体发生变化,而群体迟早也会改变它的实际环境。
杜布赞斯基(1964)指出,“在自然界存在着的各种不同的群体中,以具有两性生殖和异体受精的生物所形成的繁育集合体,最易看出群体的有机整体性。在这种孟德尔式的群体内,其整体化的因素就是生殖过程本身;这种过程把成员之间建立了交配联系、亲体联系和子代联系。所以,一个孟德尔式群体就是一个享有共同基因库(gene pool)的繁殖集团……在有性生物中,孟德尔群体(并不是个体)已成为对适应性最有决定性的自然选择形式的作用单位”。
但是,也有学者反对以利他主义为基础的群体选择。譬如,英国进化生物学家威廉姆斯(George C. Williams,1926-1010)就认为,通过利他主义的个体行为,欺诈者总是繁荣,能生存留下更多的后代,而种群中的利他主义成员将会快速灭绝,被骗子们所取代(Mills 2010)。
(4)亲属选择
群体选择中有一种称之为亲属选择(kin selection)的学说,它是由英国进化生物学家汉密尔顿(William Donald Hamilton,1936-2000)于1964年提出的。该学说认为,亲缘关系越近的动物彼此合作和利他倾向越强烈,而亲缘越远则表现越弱。典型的例子就是工蜂/工蚁(图5),它们养育·建设蜂巢/蚁巢而自己却不能繁殖后代,还要抚养女王蜂/蚁的后代。而kin selection一词则是同年由另一位英国进化生物学家和遗传学家John Maynard Smith(1920-2004)首次使用。

图5 社会昆虫蜜蜂的合作行为视为一种亲缘选择(引自维基百科)
Mills(2010)认为,汉密尔顿的亲属选择为利他主义问题提供了一个戏剧性的解答,因为亲属个体能使它们自己的基因组得到保存,这是它们乐意付出代价或自我牺牲的原因。亲属选择造成了进化论和行为的综合,即所谓的“社会生物学”(Wilson 1975)。而道金斯1976年的《自私的基因》一书被认为是将亲属选择的逻辑往前推进了一步,即归结为一个舍己为人的基因(Mills 2010)。

(5)性选择
达尔文在1871年出版的“The Descent of Man,and Selection in Relation to Sex”(中译本:人类的由来)中正式提出了“性选择”的概念。他将可能使雌雄两性根据不同生活习性通过自然选择而发生变异,或者,这一性根据另一性而发生变异的现象称之为“性选择”。
达尔文说:“这种选择的形式并不在于一种生物对于其他生物或外界条件的生存斗争上,而在于同性个体间的斗争,这通常是雄性为了占有雌性而起的斗争。其结果并不是失败的竞争者死去,而是它少留后代,或不留后代。所以性选择不如自然选择来得剧烈……任何动物的雌雄二者如果具有相同的一般生活习性,但在构造、颜色或装饰上有所不同,我相信这种差异主要是由性选择所引起的:这就是由于一些雄性个体在它们的武器、防御手段或者美观方面,比别的雄性略占优势,而这些优越性状在连续世代中又只遗传给雄性后代……在许多动物里,性选择,能够帮助普通选择保证最强健的、最适应的雄体产生最多的后代。性选择又可使雄体获得有利的性状,以与其他雄体进行斗争或对抗;这些性状将按照普遍进行的遗传形式而传给一性或雌雄两性”(Darwin 1872)。雄性孔雀艳丽的尾巴(图6)是“性选择”的经典案例。

图6 绿孔雀(Pavo muticus)(来源:维基百科)
达尔文指出,“大多数的雄性动物,如一切最美丽的鸟类,某些鱼类、爬行类和哺乳类,以及许多华丽彩色的蝴蝶,都是为着美而变得美的;但这是通过性选择所获得的成果,就是说,由于比较美的雄体曾经继续被雌体所选中……鸟类的鸣声也是这样。我们可以从一切这等情形来推论:动物界的大部分在爱好美丽的颜色和音乐的音响方面,都有相似嗜好……最简单形态的美的感觉,—即是从某种颜色、形态和声音所得到一种独特的快乐,—在人类和低于人类的动物的心理里是怎样发展起来的呢,这实在是一个很难解的问题。如果我们追究为什么某种香和味可以给与快感,而别的却给与不快感,这时我们就会遇到同样的困难”(Darwin 1872)。其实,达尔文关于动物界为何产生对美的感觉这样的疑惑迄今依然是未解之谜!
但是,精致的进攻手段、引人注目的色彩装饰和雄性求偶时炫耀行为的产生,也许有利于雄性在竞争中获胜,但这些特征由于浪费和损害了身体资源以及在性冲突中使捕猎者有机可乘,从而降低了种群的适合度(威廉斯2001)。笔者猜想,假如爱尔兰麋鹿那极度炫耀而夸张的大角是由于雌鹿畸形的审美观所致,导致雄性伴侣过度发育而终致灭绝,这该是多么悲情的性选择!
对有些性别发育可塑性大的物种来说,环境直接决定了性别的发育取向。譬如,雌雄异株的草本植物—女娄菜,当一个雌性处于雄性包围的区域中时,它的柱头将收到大量的花粉,于是它就产生过量的雌性后代(环境就是这样奖励雌性的出生),如果仅仅只有少量的花粉落在柱头上,表明本地雄性个体的稀少,这时雄性后代的比例就会增加(威廉斯2001)。不少的无脊椎动物(如蚜虫、溞和裂螠等)亦如此。
(6)其他选择
还有一些人在更高的分类层次考虑选择的对象,如物种、支序等。由于大的地貌变化导致不同生物区系的物种之间的竞争(如上新世巴拿马海峡形成后使北美和南美区系混合),引起物种的大量灭绝,称之为物种选择(但笔者认为,这更应该称为物种替换)。还有人提出支序选择理论,所谓支序指形成种系发生树上某一分支的一个分类类群,譬如白垩纪结束时整个恐龙支序灭绝了,但鸟类和哺乳动物却得以幸存。但是,迈尔(2009)认为,物种也好支序也好绝不是选择作用的对象,选择的对象仍然只能是个体。
“The theory of Evolution: What it is,Where it Came from, and Why it Works(中译本:进化论传奇—一个理论的传记)”的作者 Cynthia L. Mills女士指出,“生命支撑起了太多的现象,这些现象是不能用把它们各个组成部分的不论巨细的相加来解释的。一个生物体,并不仅仅是基因的集合,甚至也不仅是基因表达的集合。它产生于一种所谓的‘浮现’——超过各组成部分性质总合的性质的产生。生命是‘浮现’的;它不仅仅是新陈代谢反应路径和细胞结构所组成的混合环境。生命更源于自组织的结果,浮现于混沌之中”(Mills 2010)

(7)综合
Mills(2010)说,“为什么不承认选择是可以作用于基因上、个体上、群体上、亲族上,以至物种上?……如果选择在所有这些层次上发生,再结合了系统的限制和偶然性,那就再也不能靠简单、严格的法则和规律来解释进化论了”。
威廉斯(2001)说,“选择就导致了个体的产生,生命演变成为生理学上分离的个体和遗传学上独立的谱系。无源的生命就变成多源的,但是这仅发生在自然选择已经不可逆地使多肽成为代谢的工具,多聚核苷酸作为有效的信息贮存库之后。但进化成为多源的之后,它就失去了其决定论的特征,变成一种随机的过程”。
(8)没有任何选择
美国遗传学家摩尔根(Thomas Hunt Morgan,1866-1945)早期反对自然选择,赞同德弗里斯的观点,将突变作为新物种产生的直接基础。他在1903年出版的《进化和适应》一书中,不仅攻击选择机制,也攻击整个功利主义自然观,认为根本没有必要去想象选择会在任何水平起作用,任何突变只要不与环境明显不相容,就可以生存和繁殖,环境并不有效地控制进化,进化的历程完全由各种突变来决定(鲍勒1999)。
4.自然选择之我见
什么是生命延续的基本单位?当然是具有一定遗传稳定性的物种。可这样来描述生命:1)生命存在的实体是个体,2)个体的构造与生存原理源自所携带的基因组,3)物种的构造与生存原理保存在基因库之中,而基因库则分散在种群的个体之中,并通过有性生殖在世代间传递。因此,基因的目的是为了保存种族的遗传信息,服务于种族,而种族由个体组成,那么它理应服务于个体。
个体与种族利益孰重孰轻?个体,无论它采取完全孤立的生存方式还是群体合作形式,无论亲体是否照顾幼体,无论是一夫一妻还是一夫多妻(或一妻多夫),无论雌雄通过何种方式进行配对……,其行为(如本能、习性等)必定遵从有利于种族延续的内禀约束性。
自然选择什么?毫无疑问是物种,虽然延伸到了种族利益约束下的个体与基因,以及在一些类群中涉及到群体或亲属……。自然允许多样的路径或方式,只要能使物种得以延续!
从基因的视角来看,正如杜布赞斯基(1964)所说,“在实际上,选择的本质就是一个群体中的不同基因型携带者,对后代的基因库(gene pool)作出不同的贡献。在同一个环境中,有些基因型的贡献要比另一些的相对大些。一个已知基因型的携带者,把它们的基因传递到其后代基因库中去的相对能力,就是那个基因型的适应值(adaptive value),也就是达尔文的适合性(fitness)”。
个体何以能承载种族?这里并不是说个体知道为了种族的利益(直白地说,就是能够繁衍下去)而如何努力,而是说不能够成活到性成熟并成功繁衍的个体早已遭到了淘汰,剩下的个体必定具备存活与繁衍的基本技巧。不论雌雄间演绎何等崇高、或何等卑劣、何等悲催、或何等复杂、或何等平庸……之爱情故事,只要能够成功繁衍出子孙后代,自然界就会允许它的存在。
“性”是什么?就是两套DNA的抽签与组合,游戏规则就是一套来自父本,一套来自母本,从浩瀚的个体来看,这是一个随机的过程。一个物种的DNA记载了它的个体生命原则,并乘载于个(躯)体纷繁的生命之流,在延绵中变幻,在变幻中延绵!
来源:谢平. 2016. 进化理论之审读与重塑. 北京:科学出版社
(电子版下载地址:http://wetland.ihb.cas.cn/lwycbw/qt/)
作者:谢平,研究员,中国科学院水生生物研究所(xieping@ihb.ac.cn)

https://wap.sciencenet.cn/blog-1475614-1034535.html
上一篇:黑格尔的正—反—合:人类思维不断扬升的钥匙?
下一篇:论进化:坐什么井观什么天?