博文
韩国首尔大学:纳米颗粒介导肿瘤微环境中T细胞脂质代谢重编程用于免疫代谢治疗
 精选
精选
|
Nano-Micro Letters (2021)13:31 https://doi.org/10.1007/s40820-020-00555-6 2. aCD3/F/AN在葡萄糖缺乏条件下特异性激活T细胞,对肿瘤细胞产生效应杀伤作用。 3. 体内用aCD3/F/AN治疗可增加T细胞浸润、细胞因子产生,并阻止肿瘤生长。 图1. aCD3/F/ANs对T细胞的代谢重编程示意图。a. aCD3/F/ANs制备过程的示意图。b. aCD3/F/ANs促进T细胞线粒体脂肪酸代谢的机制。c. 脂肪酸代谢重编程对T细胞介导的癌细胞杀伤的影响。 图2. 纳米粒子的表征。a. ANs、ACD3/ANs、F/ANs和ACD3/F/ANs的示意图。b. 用透射电镜观察aCD3/F/ANs的形态。比例尺:200nm。c. 用DLS测定的不同纳米粒子的平均粒径。d. 用激光多普勒微电泳法测定纳米颗粒的Zeta电位。e. 通过STEM-EDS对aCD3/F/ANs的氧、氯和硫元素进行表征。比例尺:200 nm。f. 通过HPLC定量纳米粒中包裹非诺贝特的量。g. PBS中F/ANs和aCD3/F/ANs的粒径,监测7天。h. aCD3/F/ANs中非诺贝特的释放,在不同pH条件下通过HPLC定量(***P<0.001)。 II aCD3/F/Ans的T细胞摄取研究 使用荧光染料标记的纳米颗粒,通过共聚焦显微镜和流式细胞术监测T细胞对aCD3/F/ANs的摄取。aCD3/F/fANs的细胞摄取效率较F/fANs以及抗CD3抗体和fANs的混合物高。使用aCD3/F/fANs治疗后,荧光阳性T细胞数量最高,比其他治疗组高3.8倍。 图3. T细胞摄取纳米颗粒。小鼠脾源性T细胞与各种含非诺贝特的纳米颗粒制剂一起培养。24小时后,通过共聚焦显微镜观察(a)纳米颗粒的细胞摄取,并通过流式细胞仪(b、c)定量(***P<0.001,n.s.,不显著)。比例尺:10μm。 用aCD3/F/ANs处理T细胞影响脂肪酸氧化相关基因表达水平和脂质摄取。为了定量评估代谢重编程,用流式细胞仪检测PPARα在T细胞上的表达水平。在用aCD3/F/ANs处理的T细胞中,在蛋白质水平和mRNA水平上PPARα的表达最高。aCD3/F/ANs治疗组的PPARα阳性T细胞数量较未治疗组增加了7.2倍以上。此外,流式细胞术检测细胞膜上脂肪酸转位酶CD36的表达水平。aCD3/F/AN组的CD36表达较F/AN组高2.2倍。Western-blot结果显示,aCD3/F/Ans和其它组相比,显著提高了CPT1B、LCAD和MCAD的蛋白表达水平。与未处理细胞相比,CPT1B、LCAD和MCAD的mRNA水平分别增加了4.4倍、2.5倍和2.8倍。由于观察到的基因表达变化的一个结果是脂质摄取率增加,我们使用荧光染料标记的脂质BODIPYC16来测量T细胞的脂质摄取。与未经处理的细胞相比,经ANs、aCD3/ANs、F/AN或抗CD3抗体和F/ANs混合物(aCD3+F/ANs)处理的T细胞在T细胞的脂质摄取方面没有明显变化。相反,与未处理的T细胞相比,用aCD3/F/ANs处理T细胞可使脂质摄取增加3.1倍。 图4. T细胞脂肪酸代谢相关基因表达和脂质摄取。a. aCD3/F/AN诱导的脂质代谢增强的细胞机制示意图。左半部分表示未经治疗的情况,右半部分表示用aCD3/F/ANs进行治疗。采用aCD3/F/AN处理激活T细胞中的PPARα,导致脂肪酸代谢相关蛋白的过度表达,包括CD36(脂肪酸转位酶)、CPT1B、LCAD和MCAD。线粒体脂肪酸代谢的增加通过TCA循环产生更高的能量。b. 流式细胞仪测定T细胞中PPARα的相应水平(PPARα+/ CD3+)。c. 流式细胞仪检测T细胞中CD36蛋白的水平(CD36+/CD3+)。d. westernblot检测T细胞CPT1B、LCAD和MCAD蛋白表达水平。e, f. 用荧光脂质体BODIPY C₁₆孵育经不同纳米粒子处理的T细胞,流式细胞仪(e)测定荧光脂质与T细胞的结合,并用平均荧光强度(f)表示(***P<0.001)。g. RT-PCR检测脂肪酸代谢相关基因、CPT1B、LCAD和MCAD的mRNA表达水平(***P<0.001)。 IV aCD3/F/ANs激活T细胞线粒体 aCD3/F/ANs可调节T细胞的线粒体功能,包括脂肪酸代谢。与高糖条件下的T细胞线粒体相比,未经处理的T细胞或经纳米颗粒(如AN、aCD3/AN和F/AN)处理的T细胞显示出界限不清的线粒体嵴,形态缩小。与此相反,aCD3/F/ANs处理的T细胞显示线粒体结构,嵴清晰,与高糖环境下观察到的相似。aCD3/F/Ans还可以提高线粒体的膜电势。aCD3/F/AN介导的非诺贝特对T细胞的传递增强了脂质代谢和线粒体活性。 图5. T细胞线粒体形态、膜电位和脂肪酸代谢。在添加棕榈酸作为脂质来源的低葡萄糖培养基中用aCD3/F/ANs处理T细胞。a. 透射电镜显示T细胞在高糖培养基、未经aCD3/F/AN处理或经aCD3/F/AN处理的低糖培养基中的线粒体。比例尺:1μm(上面板)和500 nm(下面板)。b. 线粒体膜电位,用MitoTracker橙色CMTMRos评估,共焦荧光显微镜观察。c. 线粒体膜电位增加的T细胞群,通过流式细胞术定量。d, e. 使用Seahorse XFp分析仪测量ECAR(d)和OCR(e)。f-h. 根据ECAR和OCR值获得的Basal OCR(f)、SRC(g)和OCR/ECAR (h)值(*P<0.05,**P<0.01)。i. β‑羟丁酸分泌水平(***P<0.001)。 V 代谢重编程T细胞的体外增殖 图6. 代谢重编程诱导的T细胞存活和增殖增强。T细胞被激活并用各种纳米颗粒制剂处理,并在葡萄糖限制条件下培养,无论是否有脂质来源。a. 实验方案说明。b. 活的和死的T细胞,通过荧光染色显示。比例尺:10μm。c, d. 用annexin V和PI染色T细胞,并对各组annexin V–/PI–群体进行定量(***P<0.001)。e. 使用WST‑1分析法测量T细胞的增殖(***P<0.001)。 aCD3/F/ANs治疗增强了T细胞对癌细胞的杀伤活性。T细胞与B16F10黑色素瘤细胞共孵育后,aCD3/F/AN处理T细胞组的颗粒酶B和IFN-γ的水平分别比F/AN处理组高2.3倍和3.0倍。用aCD3/F/AN处理的T细胞孵育后死亡癌细胞的数量是用aCD3和F/Ans混合处理的的15.6倍。 图7. aCD3/F/AN处理T细胞的体外抗癌活性。用不同的纳米颗粒制剂处理T细胞,并与B16F10细胞在葡萄糖限制条件下在棕榈酸酯作为脂质源存在下共同培养。a. 实验方案说明。b, c. granzyme B (b)和IFN‑γ(c)阳性T细胞的细胞流式数据。d, e. granzyme B (d)和IFN‑γ(e)阳性T细胞群落含量。(***P<0.001)。f. 死亡癌细胞含量,流式细胞术定量(***P<0.001)。g. 实时记录通过T细胞溶解癌细胞。癌细胞用红色荧光染料标记,而T细胞用绿色荧光染料标记。 通过在荧光图像中与T细胞的共定位,可以观察到纳米颗粒在肿瘤组织中T细胞的体内分布。与F/fANs和aCD3+F/AN组相比,aCD3/F/fANs在肿瘤组织中的滞留率最高。在aCD3/F/fANs治疗组中,给药后72小时内纳米颗粒在肿瘤组织中的分布最高。纳米颗粒与CD3+T细胞的共定位在aCD3/F/fAN治疗组中最高,在24小时时表现出比F/fAN组大7.6倍。肿瘤组织染色显示aCD3/F/fANs与肿瘤内CD3+T细胞有较高的共定位。 图8. aCD3/F/ANs的体内T细胞靶向能力。通过向小鼠(n=3)瘤内注射含有非诺贝特的各种纳米颗粒制剂来评估T细胞靶向能力。a. 不同时间点的肿瘤部位的体内荧光。b. 不同时间点肿瘤部位荧光定量(**P<0.01,***P<0.001)。c. 通过流式细胞术分析肿瘤驻留T细胞中纳米粒的摄取。d. 每个治疗组定量CD3+FITC+T细胞量,(***P<0.001)。e. 共焦显微镜观察纳米颗粒(绿色)与CD3+T细胞(红色)共定位。比例尺:20μm。 肿瘤内注射aCD3/F/Ans可以通过T细胞调控体内的PPARα的表达、脂质摄取和代谢。流式细胞仪检测PPARα‑阳性和BODIPY C₁₆阳性。aCD3/F/AN治疗组的T细胞数量增加的幅度更大。aCD3/F/AN治疗组的T细胞数量增加的幅度更大。与F/AN和aCD3+F/AN治疗组相比,aCD3/F/AN治疗组PPARα的表达分别增加了4.1倍和2.6倍。aCD3/F/AN治疗组的BODIPY C₁₆脂质摄取几乎是其他组的两倍。与未治疗组相比,aCD3/F/AN治疗组小鼠CPT1B、LCAD和MCAD的蛋白表达和mRNA水平显著增加。免疫组化显示在肿瘤组织中CD3+T细胞中不同的PPARα表达。MALDI成像显示用aCD3/F/ANs处理的T细胞,其脂肪酸氧化代谢产物乙酰乙酸,β‑羟丁酸和棕榈酰肉碱有所增强。 图9. 体内肿瘤组织中T细胞的PPARα表达和脂质吸收。a. B16F10荷瘤小鼠在肿瘤接种后第10天和第12天瘤内注射各种纳米颗粒制剂。第13天,瘤内注射BODIPY C16,第14天,提取并分析肿瘤组织。b-e. 通过流式细胞术分析T细胞(b,d)和BODIPY C16阳性CD3 T细胞(c,e)的PPARα表达水平(***P<0.001)。f. westernblot检测肿瘤浸润T细胞CPT1B、LCAD和MCAD蛋白表达水平。g. 通过RT-PCR(***P<0.001)测定肿瘤浸润T细胞中脂肪酸代谢相关基因CPT1B、LCAD和MCAD的mRNA表达水平。h, i. 未治疗组(h)和aCD3/F/AN治疗组(i)的肿瘤组织的共聚焦显微镜图像用抗CD3(绿色)和抗PPAR进行免疫染色α (红色)抗体和DAPI复染。比例尺:100μm。j. β‑MALDI成像显示乙酰乙酸(m/z=103.09),羟丁酸(m/z=105.10)和棕榈糖基肉碱(m/z=400.60)的肿瘤组织分布。比例尺:5 μm。 为评价aCD3/F/ANs对小鼠肿瘤生长和存活的影响,我们在B16F10荷瘤小鼠瘤内注射两次,接种后第10天注射一次,第12天再次注射不同的纳米颗粒制剂。结果显示,aCD3/F/AN治疗的肿瘤生长抑制效果显著高于其他纳米颗粒制剂。为了验证观察到的抗肿瘤疗效是否归因于代谢重编程T细胞的功能增强,我们对肿瘤浸润性CD8+T细胞进行了定量。结果显示,在用aCD3/F/ANs治疗的小鼠中,肿瘤组织中的CD8+T细胞数量显著增加,肿瘤组织中的CD8+T细胞数量是F/AN治疗组小鼠的14.8倍。aCD3/F/ANs的体内治疗影响IFN-α的表达水平γ T细胞颗粒酶B。aCD3/F/AN治疗组中表达的干扰素-γ肿瘤T细胞占20.5%±4.1%,比F/AN和aCD3+F/AN治疗组增加了5倍以上。与干扰素γ的表达模式类似, aCD3/F/ANs组表达granzyme B的T细胞百分比最高,比其他治疗组表达水平增加了3.9倍以上。对肿瘤组织的免疫组化分析显示,未治疗小鼠的肿瘤组织中没有明显的CD8+T细胞浸润。然而,在用aCD3/F/ANs治疗组的肿瘤组织中可以观察到有大量CD8+T细胞浸润和IFN-γ和颗粒酶B的分泌。 图10. aCD3/F/ANs的体内抗肿瘤作用。a. B16F10荷瘤小鼠在肿瘤接种后第10天和第12天瘤内注射不同的纳米颗粒制剂。b, c. 通过测量肿瘤体积来确定抗肿瘤疗效。d.各组小鼠存活率,肿瘤接种后60天监测。 本文通讯作者 Seoul National University 多功能纳米生物材料递送化学和核酸药物。 ▍主要研究成果 ▍Email: ohyk@snu.ac.kr Nano-Micro Letters《纳微快报(英文)》是上海交通大学主办、Springer Nature合作开放获取(open-access)出版的英文学术期刊,主要报道纳米/微米尺度相关的高水平文章(research article, review, communication, commentary, perspective, letter, highlight, news, etc),包括微纳米材料的合成表征与性能及其在能源、催化、环境、传感、吸波、生物医学等领域的应用研究。已被SCI、EI、SCOPUS、DOAJ、CNKI、CSCD、知网、万方、维普等数据库收录。2020 JCR影响因子:12.264。在物理、材料、纳米三个领域均居Q1区(前10%)。2020 CiteScore:12.9,材料学科领域排名第4 (4/120)。中科院期刊分区:材料科学1区TOP期刊。全文免费下载阅读(http://springer.com/40820),欢迎关注和投稿。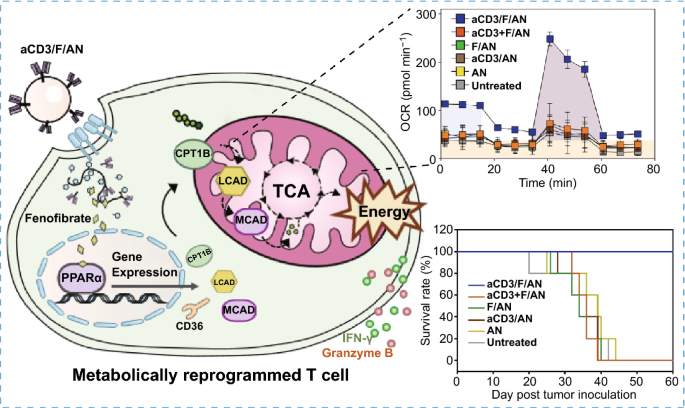
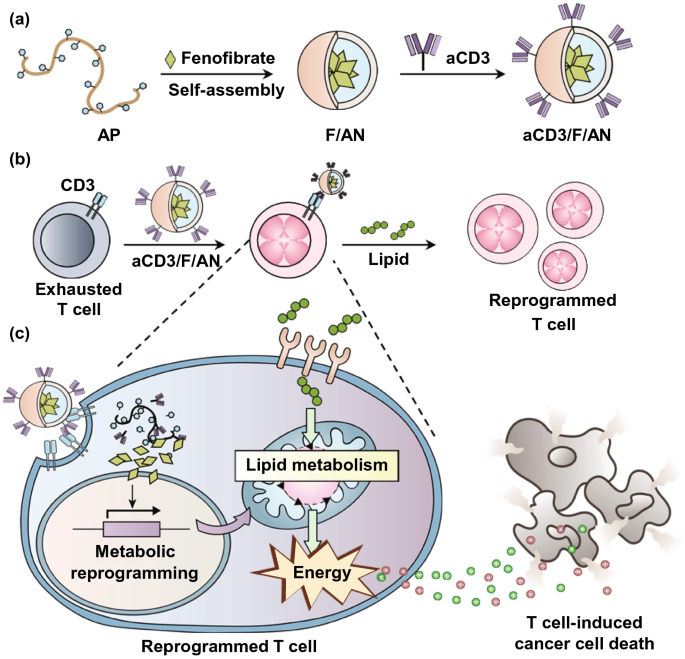
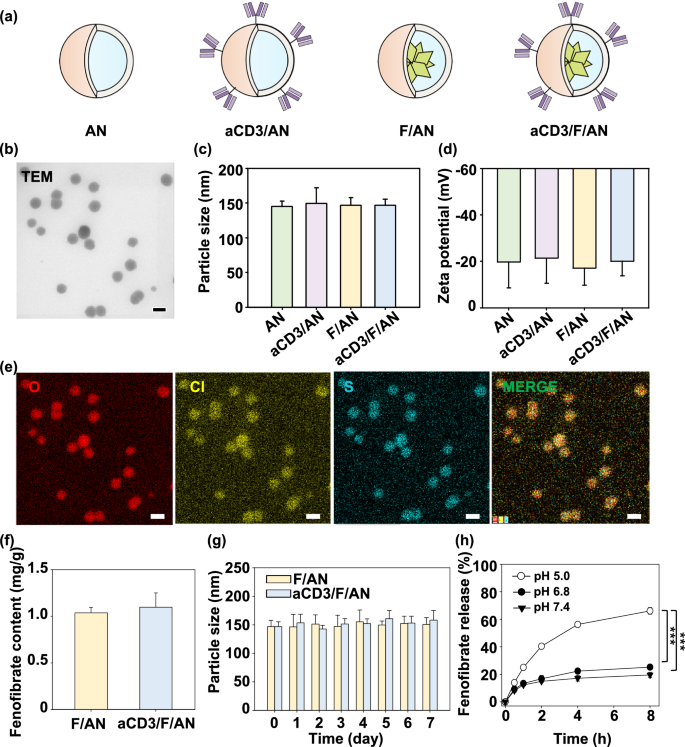
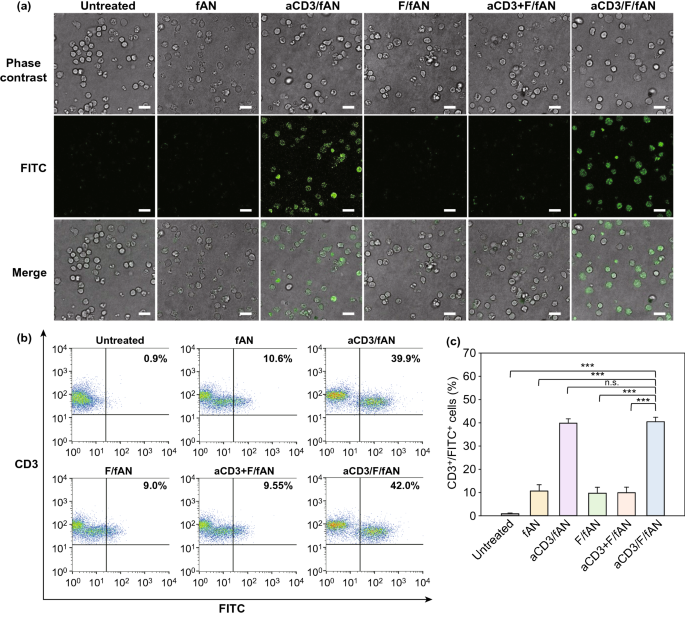
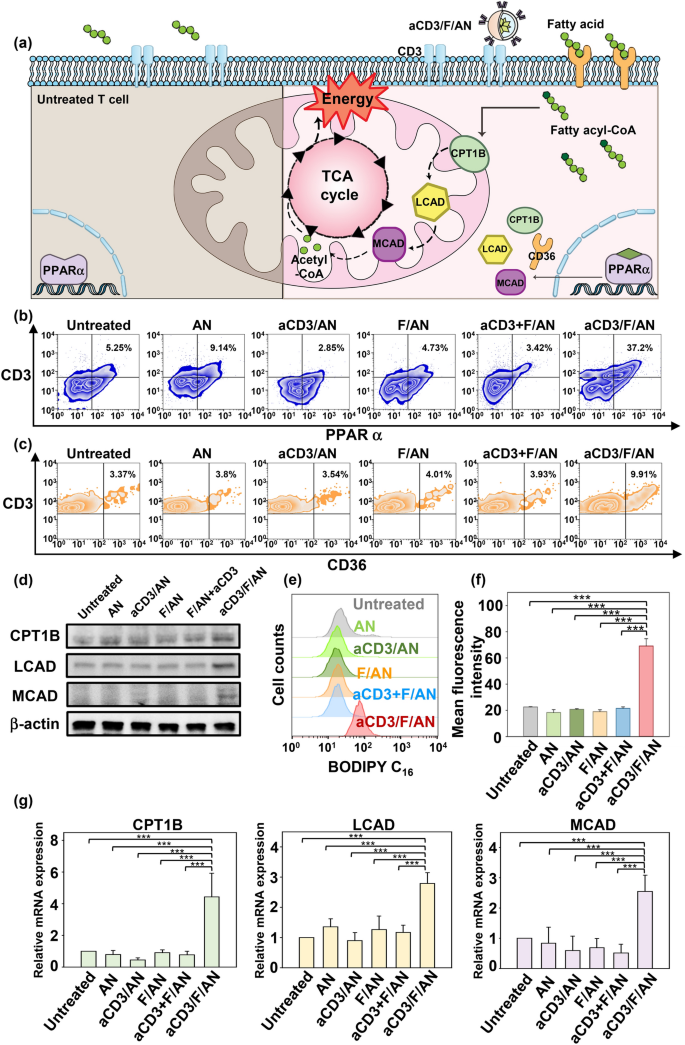
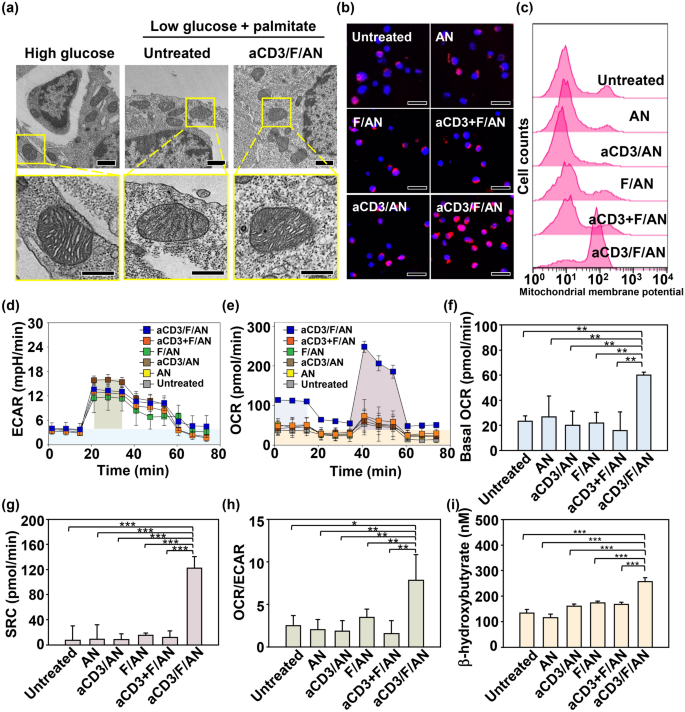
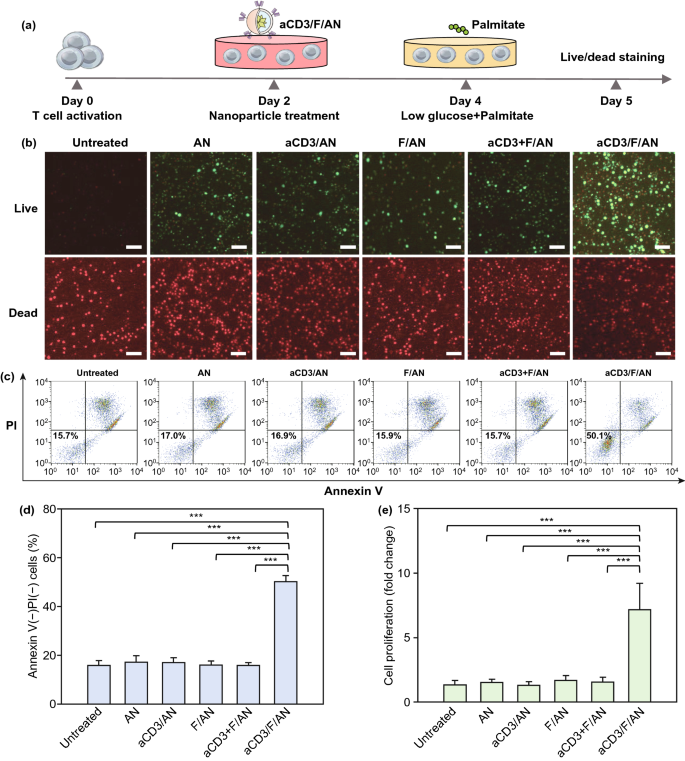
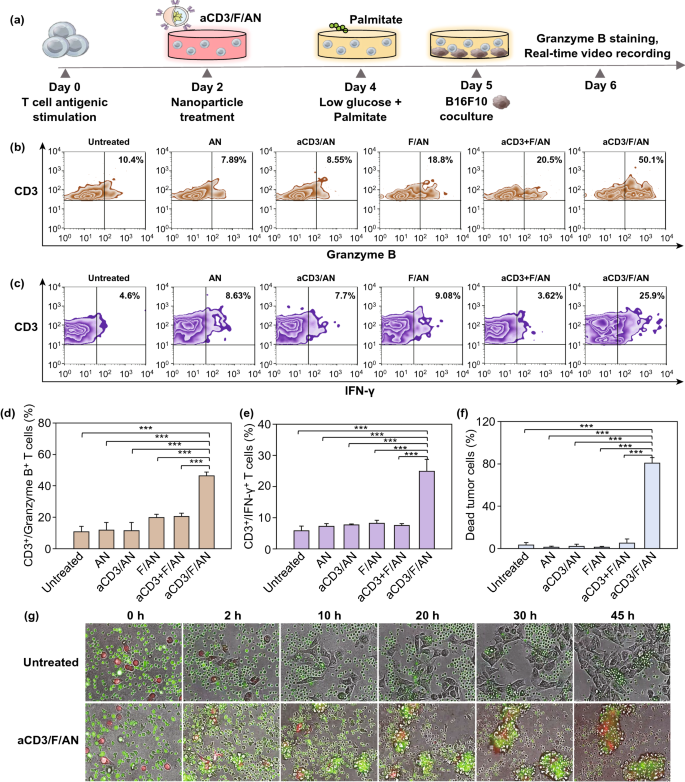
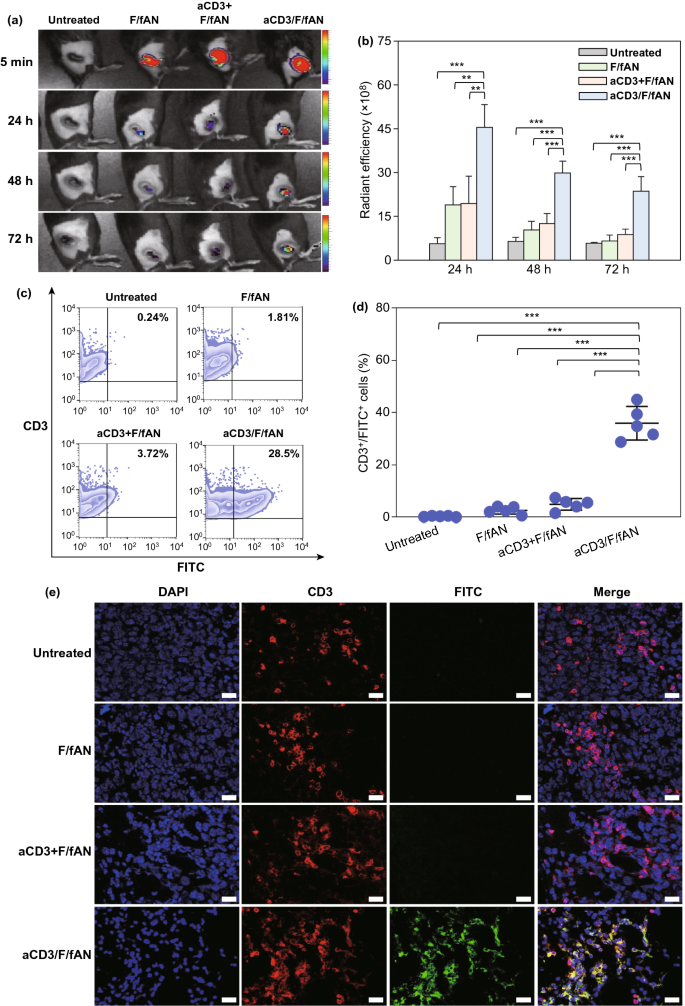
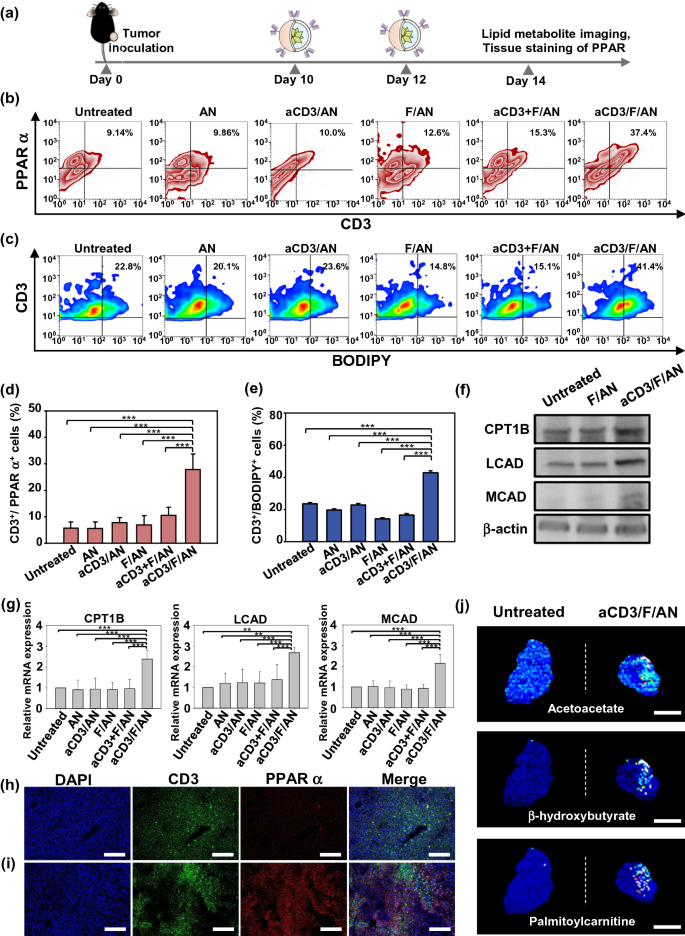
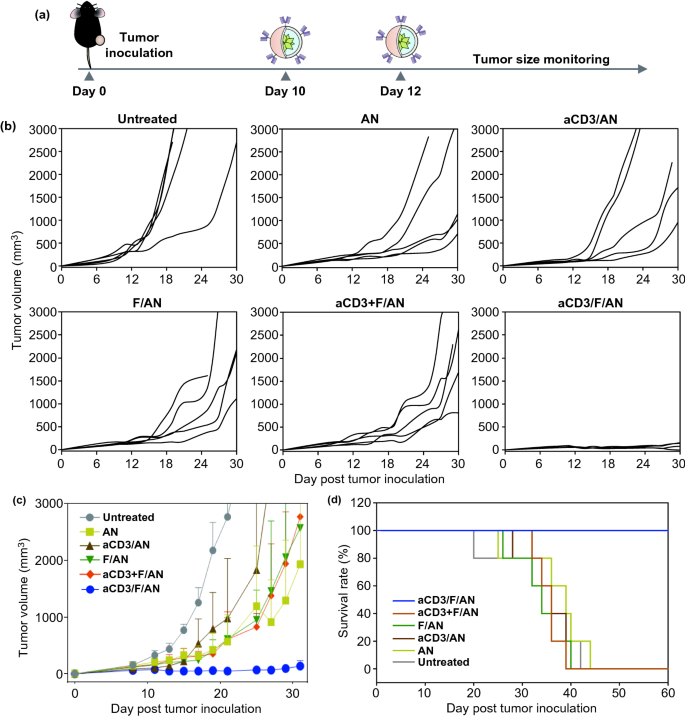
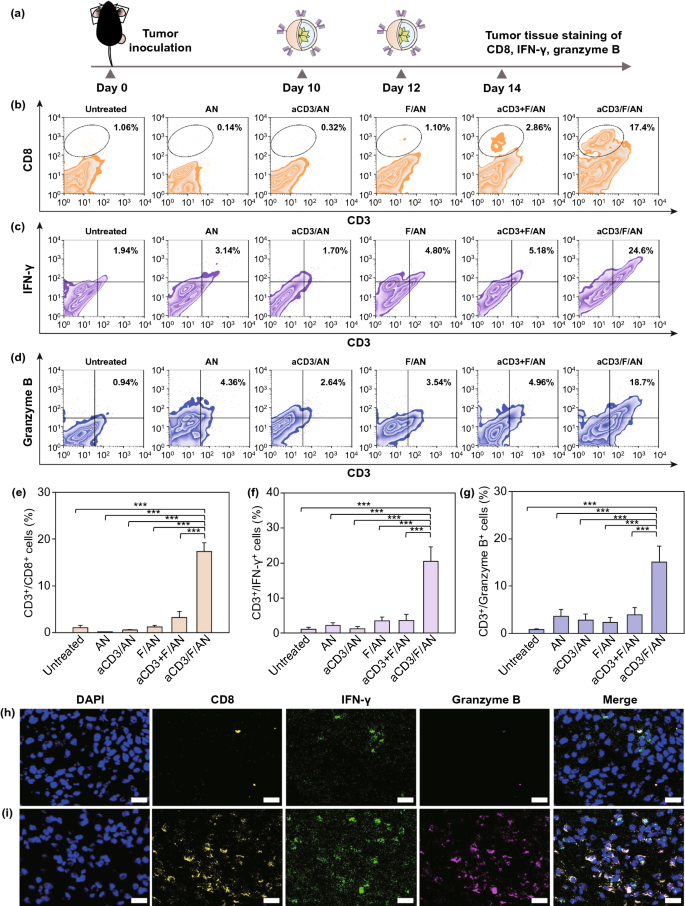
图11. T细胞在肿瘤组织中产生细胞因子。a. B16F10荷瘤小鼠在肿瘤接种后第10天和第12天注射纳米颗粒制剂。第14天,提取肿瘤组织进行分析。b-d. 流式细胞术检测浸润性CD8+T细胞(b),肿瘤组织CD3+T细胞中表达干扰素-γ的T细胞(c)和颗粒酶B的T细胞(d)(**P<0.001)。CD3+/CD8+细胞群(e),CD3+/IFN‑γ+细胞(f)和CD3+/颗粒酶+细胞(g)。h, i. 在未治疗(h)和aCD3/F/AN治疗(i)的小鼠中,提取肿瘤组织并用抗CD8(黄色)、抗IFN-γ进行免疫染色(绿色)和抗颗粒酶B(浅紫色)抗体,并用DAPI复染。比例尺:20μm。 Yu‑Kyoung Oh
Yu‑Kyoung Oh
https://wap.sciencenet.cn/blog-3411509-1290760.html
上一篇:日本金泽大学等:TiO₂多层前端接触提升钙钛矿太阳能电池效率
下一篇:封面文章:基于独立式纳米传感器的远程气体监测系统